

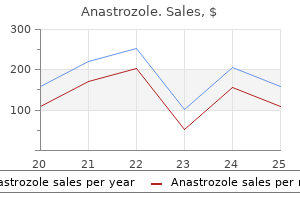
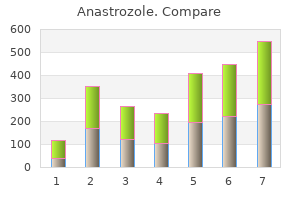
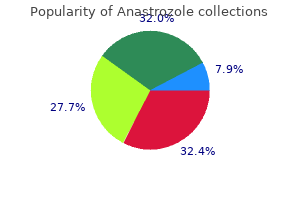
Anastrozole
| Contato
Página Inicial
Teresa M. Bieker, MBA-H, RDMS, RDCS, RVT
- Lead Sonographer
- Division of Ultrasound
- University of Colorado Hospital
- Aurora, Colorado
The anterior part of the ventrolateral nucleus projects to the premotor and supplementary motor areas of cortex with no projection to area 4 women's health center medina ny cheap 1 mg anastrozole visa. The projection to the motor cortex arises in areas 1 and 2 women's health center naperville il anastrozole 1 mg otc, with little or no contribution from area 3b womens health zone link health 1 mg anastrozole with amex. It has been suggested that this pathway plays a role primarily in making motor adjustments during a movement pregnancy week 6 cheap anastrozole 1 mg without prescription. Neurones in area 4 are responsive to peripheral stimulation menopause joint aches order anastrozole canada, and have receptive fields similar to those in the primary sensory cortex. Cells located posteriorly in the motor cortex have cutaneous receptive fields, whereas more anteriorly situated neurones respond to stimulation of deep tissues. The motor cortex receives major frontal lobe association fibres from the premotor cortex and the supplementary motor area, and also fibres from the insula. It is probable that these pathways modulate motor cortical activity in relation to the preparation, guidance and temporal organization of movements. Area 4 sends fibres to , and receives fibres from, its contralateral counterpart, and also projects to the contralateral supplementary motor cortex. Apart from its contribution to the corticospinal tract, the motor cortex has diverse subcortical projections. The motor cortex sends projections to all nuclei in the brainstem, which are themselves the origin of descending pathways to the spinal cord: namely, the reticular formation, the red nucleus, the superior colliculus, the vestibular nuclei and the inferior olivary nucleus. It can either be situated completely over the lateral fissure or be in part internal to the fissure, in this situation giving the false impression that the central sulcus is a branch of the lateral fissure. The superior connection corresponds to the paracentral lobule (of Ecker) disposed along the medial surface of the hemisphere inside the interhemispheric fissure, delineated anteriorly by the paracentral sulcus and posteriorly by the ascending and distal part (marginal ramus) of the cingulate sulcus. Broca described a middle connection between the pre- and postcentral gyri (pli de passage moyen of Broca) that may be present as a gyral bridge, usually hidden within the central sulcus; on the cortical surface, this corresponds to the classic, posteriorly convex, middle genu of the central sulcus. When this middle connection is sufficiently developed so that it reaches the brain surface, it interrupts the central sulcus (Régis et al 2005). The localization of motor and sensory hand areas has been studied by correlating imaging cortical stimulation and postmortem cadaveric studies. Postmortem studies revealed that this protrusion was delimited by two anteriorly directed fissures that deepened towards the base of the protrusion. Hand sensory function has been localized to the postcentral component of the middle connection of the pre- and postcentral gyri (Boling and Olivier 2004, Boling et al 2008). The precentral gyrus is delimited anteriorly by the precentral sulcus, itself divided into superior and inferior precentral sulci by the connection of the middle frontal gyrus with the precentral gyrus. Further connections of the superior, middle and inferior frontal gyri may divide the superior and the inferior precentral sulci into additional segments. More dorsally, within the precentral region, the marginal precentral sulcus (sulcus precentralis marginalis of Cunningham) may merge with the superior precentral or central sulci. The inferior segment of the precentral sulcus always ends inside the opercular part of the inferior frontal gyrus, producing its characteristic U shape. The superior frontal gyrus is continuous anteriorly and inferiorly with the rectus gyrus; it may also be connected to the orbital gyri and the middle frontal gyrus. Posteriorly, it is connected to the precentral gyrus by at least one fold, which most commonly lies medially along the interhemispheric fissure. Usually the superior longitudinal gyrus is subdivided into two longitudinal portions by a medial frontal sulcus; its medial portion is sometimes termed the medial frontal gyrus. The supplementary motor area is located along the most medial portion of the superior frontal gyrus, immediately facing the precentral gyrus; it varies between individuals and has poorly defined borders. The middle frontal gyrus is usually the largest of the frontal gyri, frequently connected superficially to the precentral gyrus by a prominent root that lies between the extremities of a marked interruption in the precentral sulcus. It harbours a complex of multiple shallow sulcal segments known collectively as the middle or intermediate frontal sulcus (Petrides 2012). Superiorly, the inferior frontal gyrus is crossed by various small branches of the interrupted inferior frontal sulcus; the triangular sulcus typically pierces the superior aspect of the triangular part. The most posterior aspect of the inferior frontal gyrus, identifiable by the connection of its opercular part with the precentral gyrus, corresponds to the ventral premotor cortical area; its bilateral stimulation causes speech arrest (Duffau 2011). It is very deep and is frequently continuous, ending posteriorly by encroaching on the precentral gyrus at the level of its omega region (corresponding to the motor cortical representation of the contralateral hand). The superior frontal sulcus therefore tends to point the way to the middle frontoparietal pli de passage, as well as to the middle genu of the precentral gyrus, where there is also a motor representation of the hand (Boling et al 1999). The inferior frontal sulcus is always interrupted by the multiple connections running between the middle and inferior gyri and usually has three parts: orbital, triangular and opercular. The triangular part is usually more retracted, such that there is a small widening of the lateral fissure at its base corresponding to the anterior Sylvian point. It is characterized by horizontal and anterior Cerebral hemispheres ascending rami of the lateral fissure that consistently divide the lateral fissure into anterior and posterior branches. The anterior basal portion of the opercular part is sometimes divided by another branch of the lateral fissure, the diagonal sulcus of Eberstaller. Inferiorly, the orbital part continues with the lateral orbital gyrus, at times passing under a shallow sulcus known as the fronto-orbital sulcus. The basal apex of the triangular part is always superior to the lateral fissure; the base of the opercular part can be located either superiorly or within the fissure. Anteriorly, the inferior frontal gyrus terminates by merging with the anterior portion of the middle frontal gyrus. All of the frontal gyri are delineated anteriorly by the frontomarginal sulcus (frontomarginal sulcus of Wernicke), which lies superior and parallel to the supraciliary margin, separating the superolateral and orbital frontal surfaces. Posteriorly, the inferior frontal gyrus is connected to the precentral gyrus along the posterior aspect of its opercular part. The olfactory sulcus lies longitudinally in a paramedian position on the frontobasal or orbital surface of each frontal lobe. The narrow gyrus rectus, medial to the olfactory sulcus, is considered to be the most anatomically constant of the cerebral gyri. The orbital gyri, lateral to the olfactory sulcus, account for the greatest proportion of the frontobasal surface. The anterior, posterior, medial and lateral orbital gyri are delineated by the lateral, medial and transverse orbital sulci and the cruciform sulcus of Rolando, which together form a characteristic H shape. The posterior orbital gyrus lies anterior to the anterior perforated substance and typically presents a configuration similar to a tricorn hat, a feature that may facilitate its identification in anatomical specimens where the H-shaped orbital sulcus is less obvious. The remaining orbital gyri are connected to the superior, middle and inferior frontal gyri along the frontal pole. Anterior to the paracentral lobule, the medial aspect of the superior frontal gyrus lies over the cingulate sulcus and the cingulate gyrus, merging inferiorly with the gyrus rectus. The latter is bounded superiorly by the superior rostral sulcus and accommodates the shallower inferior rostral sulcus along its surface. The cingulate gyrus systematically connects with the gyrus rectus around the posterior end of the superior rostral sulcus by a prominent U-shaped cortical fold known as the cingulate pole, which is located immediately anterior to the subcallosal gyri. Small supraorbital sulci lie within the medial surface of the frontal pole, superior to the superior rostral sulcus at the level of the genu of the corpus callosum. The most distal component of the operculum is a C-shaped convolution that connects the supramarginal and superior temporal gyri, and encircles the posterior end of the lateral (Sylvian) fissure. The bases of the U-shaped convolutions and their related sulcal extremities may be either superior to the fissure, as indicated in this specimen, or inside the fissure. A homologous pathway to the brainstem, the corticonuclear projection, fulfils a similar function in relation to motor nuclei of the brainstem (Ch. The percentage of corticospinal fibres that arise from the primary motor cortex may be in the region of 2030%. They arise from pyramidal cells in layer V and give rise to the largest-diameter corticospinal axons. There is also a widespread origin from other parts of the frontal lobe, including the premotor cortex and the supplementary motor area. Many axons from the frontal cortex, notably the motor cortex, termi- nate in the ventral horn of the spinal cord. In cord segments mediating dexterous hand and finger movements, they terminate in the lateral part of the ventral horn, in close relationship to motor neuronal groups. The majority of parietal fibres to the spinal cord terminate in the deeper layers of the dorsal horn. The demarcation of the temporal, parietal and occipital lobes according to two different systems is shown. In one system, a parietotemporal line is drawn from the lateral edge of the parieto-occipital sulcus (2) to the preoccipital notch (temporo-occipital incisure) (4). This line sets the arbitrary anterior border of the occipital lobe (O), separating it from the parietal and temporal lobes anterior to it. A second arbitrary temporo-occipital line (5) is drawn from the posterior descending ramus of the Sylvian fissure (3) to the middle of the parietotemporal line (6). The central sulcus (1) usually curves onto the medial surface perpendicular to the marginal segment of the cingulate sulcus. A line drawn from the central sulcus to the cingulate sulcus establishes the frontoparietal border. The deep parieto-occipital sulcus (2) demarcates the parietal lobe from the occipital lobe. An arbitrary basal parietotemporal line (8) drawn from the inferior end of the parieto-occipital sulcus to the preoccipital notch establishes the temporal (T)/occipital (O) border. The limbic lobe (L) is delimited by the cingulate sulcus (9), the subparietal sulcus (10) and the collateral sulcus (11). B, the sensory homunculus showing proportional somatotopical representation in the somaesthetic cortex. Their activity precedes the onset of electromyographic activity by 50100 milliseconds, suggesting a role for cortical activation in generating rather than monitoring movement. It extends on to the medial surface, where it becomes contiguous with area 24 in the cingulate gyrus, anterior and inferior to the paracentral lobule. Lateral area 6, the area over most of the lateral surface of the hemisphere, corresponds to the premotor cortex. The major thalamic connections of the premotor cortex are with the anterior division of the ventrolateral nucleus and with the centromedian, parafascicular and centrolateral components of the intralaminar nuclei. Subcortical projections to the striatum and pontine nuclei are prominent, and this area also projects to the superior colliculus and the reticular formation. Commissural connections are with the contralateral premotor, motor and superior parietal (area 5) cortex. Ipsilateral corticocortical connections with area 5 in the superior parietal cortex, and inferior parietal area 7b, are common to both dorsal and ventral subdivisions of the premotor cortex, and both send a major projection to the primary motor cortex. The dorsal premotor area also receives fibres from the posterior superior temporal cortex and projects to the supplementary motor cortex. The supplementary motor cortex receives connections from widespread regions of the ipsilateral frontal lobe, including from the primary motor cortex, the dorsal premotor area, the dorsolateral and ventrolateral prefrontal, medial prefrontal and orbitofrontal cortex, and the frontal eye field. These connections are reciprocal but the major ipsilateral efferent pathway is to the motor cortex. Parietal lobe connections of the supplementary motor cortex are with the superior parietal area 5 and possibly inferior parietal area 7b. Contralateral connections are with the supplementary motor area, and motor and premotor cortices of the contralateral hemisphere. Subcortical connections, other than with the thalamus, pass to the striatum, subthalamic nucleus and pontine nuclei, the brainstem reticular formation and the inferior olivary nucleus. The supplementary motor area makes a substantial contribution to the corticospinal tract, contributing as much as 40% of the fibres from the frontal lobe. The supplementary motor area contains a representation of the body in which the leg is posterior and the face anterior, with the upper limb between them. Its role in the control of movement is primarily in complex tasks that require temporal organization of sequential movements and in the retrieval of motor memory. Stimulation of the supplementary motor area in conscious patients has been reported to elicit the sensation of an urge to move, or of anticipation that a movement is about to occur. The perimeter of these areas is delineated by an interrupted line to indicate uncertainty as to their precise extent. Areas 22 and 37 are considered by some to be respectively auditory and visuo-auditory areas associated with speech and language. All of these association connections are likely to be, or are known to be, reciprocal. Neuronal activity in the premotor cortex in relation to both preparation for movement and movement itself has been extensively studied experimentally. In behavioural tasks, neurones in the dorsal premotor cortex show anticipatory activity and task-related discharge as well as direction selectivity, but little or no stimulus-related changes. The dorsal premotor cortex is probably important in establishing a motor set or intention, contributing to motor preparation in relation to internally guided movement. In contrast, ventral premotor cortex is more related to the execution of externally (especially visually) guided movements in relation to a specific external stimulus. It receives its major thalamic projection from the parvocellular mediodorsal nucleus, with additional afferents from the medial pulvinar, the ventral anterior nucleus and the suprageniculate limitans complex, and connects with the paracentral nucleus of the intralaminar group. The thalamocortical pathways to the frontal eye field form part of a pathway from the superior colliculus, the substantia nigra and the dentate nucleus of the cerebellum. The frontal eye field has extensive ipsilateral corticocortical connections, receiving fibres from several visual areas in the occipital, parietal and temporal lobes, including the medial temporal area (V5) and area 7a. There is also a projection from the superior temporal gyrus, which is auditory rather than visual in function. From within the frontal lobe, the frontal eye field receives fibres from the ventrolateral and dorsolateral prefrontal cortices. It projects to the dorsal and ventral premotor cortices and to the medial motor area, probably to the supplementary eye field adjacent to the supplementary motor area proper.
Their subsequent development varies according to the type of joint that is formed women's health center lagrange ga buy 1 mg anastrozole with mastercard. In developing fibrous joints women's health clinic jersey city purchase generic anastrozole, the interzone is converted into collagen breast cancer in males order anastrozole 1 mg visa, which is the definitive medium connecting the bones involved pregnancy jaw pain buy anastrozole online now. In developing synchondroses menstruation 3 days late anastrozole 1 mg buy on-line, the interzone becomes (growth) cartilage of the modified hyaline type; in developing symphyses, it is predomi nantly fibrocartilage. In developing synovial joints, the interzonal mes enchyme becomes trilaminar when a more tenuous intermediate zone appears, splitting the mesenchyme into two dense strata. As the skeletal elements chondrify, and in part ossify, the dense strata of the interzonal mesenchyme also become cartilaginous; subsequent cavitation of the intermediate zone establishes the cavity of the joint. The loose mesen chyme around the cavity forms the synovial membrane and probably also gives rise to all other intraarticular structures, such as ligaments, discs and menisci. In joints containing discs or menisci, and in com pound articulations, more than one cavity may appear initially, some times merging later into a complex single cavity. As development proceeds, thickenings in the fibrous capsule can be recognized as the specializations peculiar to a particular joint. The columels consist of groups of individual pools of differentiated motor neurones, precisely organized according to function (Jessell et al 2011). Neurones in medial pools express the transcription factor Islet1 and their axons innervate ventral limb muscles, while neurones in lateral pools express the related transcrip tion factor, Lim1, and innervate dorsal limb muscles. Neurones also express different Hox proteins according to their position along the rostrocaudal axis of the spinal cord, and this determines their identity and their position within the spinal cord (Dasen et al 2005). Motor axons enter the developing limbs earlier than sensory axons at the time when dorsal and ventral muscle masses have started to form. Guidance of the growing axons is not dependent on the presence of migrating myoblasts; when these are absent, the main nerve trunks in the limb still form, although the smaller motor branches that would innervate individual muscles do not form (Lewis et al 1981, Honig et al 2005). Motor axons grow into the dorsal and ventral muscle masses and innervate muscle groups in a proximodistal progression. Experimental studies in mice and chickens have shown that axons of neurones in lateral pools grow into the dorsal part of the limb as a result of repulsive interactions between the receptor EphA4 (expressed by these motor neurones) and its ligand, ephrinA (expressed by ventral limb cells). The axons of EphBexpressing motor neurones in medial pools grow into the ventral part of the limb as a result of repulsive interactions with ephrinBexpressing cells in the dorsal limb (Drescher 2010). These cells are committed at an early stage and can be identified in the lateral halves of the somites. Cells from the dorsomedial edge of the dermomyotomes form the axial musculature, whereas, at limb levels, cells deepithelialize and migrate from the ventrolateral edges of the dermomyotome into the limb bud. These precursor limb myoblasts migrate through a non random, structured network of extracellular fibrils. At their leading ends, the migrating cells exhibit filopodia, which are in contact with the extracellular fibrils or with other cells; it is believed that the orienta tion of the extracellular fibrils may direct the migration of the cells. The precursor muscle cells do not differentiate into muscle before their 222 vasculature of the limb dorsoventral axis of the limb and that results in dorsal expression of Lmxb1. Sensory neurones in the dorsal root ganglia opposite the limbs send axons peripherally to terminate in the skin and to innervate propriocep tors in the limb muscles. The trajectories of sensory axons into the limb can be influenced by interaction with motor axons. When sensory axons grow into the developing limb, it has already been invaded by motor axons and the environment will have been altered by any extracellular factors they may have secreted. If motor axons are removed before sensory axons extend into the limb, those sensory axons, which would normally terminate in muscle (Ia afferent axons), appear unable to do so and instead become cutaneous nerves (Honig et al 1986, Honig and Rutishauser 1996, Honig et al 1998). The developing skin (ectoderm and immediately underlying somato pleuric mesenchyme) is essential for the normal development of cuta neous sensory axons. If the ectoderm is removed, the cutaneous nerves that would project to it are absent. Cutaneous sensory axons are not matched to particular regions of skin in the limb and can innervate regions outside their usual anatomical dermatomes, or expand into regions of skin where the cutaneous inner vation is eliminated (Wang and Scott 2002). The central processes of the proprioceptive sensory neurones innervating limb muscle form mono synaptic connections to motor neurones in the spinal cord after their peripheral axons have extended into the limbs. However, the pattern for these sensory/motor connections appears to be specified early. The proximodistal position of the sensory endings in the limb is linked to the dorsoventral location of the target motor neurones in the spinal cord, such that proximal sensory axons will connect to ventral motor columels (Sürmeli et al 2011). The different types of sensory endings show activation at different developmental times; mechanoreceptors and proprioceptors are active ahead of nociceptive neurones prenatally. A third wave of mechanosen sitivity acquisition by the remaining nociceptors occurs just after birth (Lechner et al 2009). This restriction of normal fetal movements in late gestation reflects the degree of compliance of the maternal uterus; there is a slowing of growth at this time. In addition to promoting normal musculoskeletal development, movements of the fetus encourage skin growth and flexibility indirectly. Fetuses with in utero muscular dystrophies, or other conditions that result in small or atrophied muscles, have webs of skin, pterygia, which pass across the flexor aspect of the joints and severely limit movement. Multiple pterygium syndrome is characterized by webbing across the neck, the axillae and antecubital fossae. Usually, the legs are maintained straight and webbing is not seen at the hip and knee. These conditions may be recog nized on prenatal ultrasound examination by the appearance of fixed, immobile limbs in bizarre positions, or by webbing in limb flexures. Specific syndromes, lethal multiple pterygium syndrome and congenital muscular dystrophy, have been described. The workload undertaken by the musculoskeletal system before birth is relatively light because the fetus is supported by the amniotic fluid and, therefore, under essentially weightless conditions. The load on the muscles and bones is generated by the fetus itself, with little gravitational effect. The reduction of gravitational force afforded by the supporting fluid means that all parts of the fetus are subject to relatively equal forces and that the position assumed by the fetus relative to gravity is of little consequence. This is important to ensure the normal modelling of fetal bones, especially the skull. Skulls of premature babies may become distorted as a result of the weight of the head on the mattress, despite regular changes in position, and the application of oxygen therapy via a mask attached by a band around the head can cause dysostosis of the occipital bone. Later, one main vessel, the axial artery, supplies the limb and the terminal plexus. The development of the vasculature in the limb precedes the mor phological and molecular changes that occur within the limb mesen chyme as tissues begin to form. Cartilage differentiation within the chick limb bud occurs only after local vascular regression begins, and only in areas with few or no capillaries (Hallmann et al 1987). It is not known whether the presence, or lack, of blood vessels affords different local environmental stimuli for mesenchymal cells (by varying the supply of nutrients to the tissue), or whether the local environment is controlled by the endothelial cells. Similarly, it is not clear whether inductive factors from the limb mesenchyme cause the changes that occur in the pattern of blood vessels. Work on chick wing buds suggests that the position of the central artery in the primitive limb bud vascu lature depends on Shh signalling from the polarizing region (Davey et al 2007). Simple movements of an extremity have been observed sporadically as early as the seventh week of gestation in human embryos. Combined movements of limb, trunk and head commence between 12 and 16 weeks of gestation. Movements of the embryo and fetus encourage normal skin growth and flexibility, in addition to the progressive maturation of the muscu loskeletal system. Movements of the fetus often involve slow and asym metric twisting and stretching movements of the trunk and limbs, which resemble athetoid movements. There may also be rapid, repeti tive, wideamplitude limb movements, similar to myoclonus. By term, the quality of the movements has generally matured to smooth, alternating movement of the limbs, with medium speed and intensity. The reduced effect of gravity in utero may cause certain fetal movements to appear, on ultrasonography, more fluent than the equiva lent movements observed postnatally. As the limb enlarges, the marginal vein can be subdivided into pre and postaxial veins, which run along their respective borders and which are the precursors of the superficial veins of the limb. Generally, the preaxial (superficial) veins join the deep veins at the proximal joint, and the postaxial (superficial) veins join the deep veins at the distal joint of the limb. In: Ferretti P, Copp A, Tickle C et al (eds) Embryos, Genes and Birth Defects, 2nd ed. Zeller R, LopezRios J, Zuniga A 2009 Vertebrate limb development; moving towards integrative analysis of organogenesis. Chevallier A, Kieny M, Mauger A 1977 Limbsomite relationship: origin of the limb musculature. Lewis J, Chevallier A, Kieny M et al 1981 Muscle nerve branches do not develop in chick wings devoid of muscle. An account that reviews and contrasts recent work on both chick and mouse models on the importance of movement in the development of the locomotor apparatus. A review of recent work using chicken embryos to explore the importance of movement in development of the locomotor apparatus. Sugimoto Y, Takimoto A, Akiyama H et al 2013 Scx+/Sox9+ progenitors contribute to the establishment of the junction between cartilage and tendon/ligament. Summerbell D, Lewis J, Wolpert L 1973 Positional information in chick limb morphogenesis. Sürmeli G, Akay T, Ippolito G et al 2011 Patterns of spinal sensorymotor connectivity prescribed by a dorsoventral positional template. Weintraub H, Davis R, Tapscott S et al 1991 the MyoD gene family: nodal point during specification of the muscle cell lineage. A review of the molecular basis of limb development particularly focusing on how cellcell interactions are integrated. Zuniga A, Zeller R, Probst S 2012 the molecular basis of human congenital limb malformations. A presentation of recently identified genes involved in limb development and their relevance to human congenital limb defects. This effort started with the formalizations of human, fruit fly and mouse anatomy in the 1990s and now includes all the major model organisms (Bard 2005, Druzinsky et al 2013). The types of data that are associated with tissues currently include gene expression, diseases and abnormal phenotypes. Although most users do not need to know in detail how anatomy is handled within a computational context, there are two key aspects that should be appreciated. Part of, in particular, has several meanings and these may or may not need to be distinguished. The great advantage of this approach is that the triples can be easily linked (the interventricular septum is part of the heart is part of the cardiovascular system) to make a hierarchy or network (more formally, this is a mathematical graph made of nodes and edges). These relationships enable specific queries about the knowledge included in the ontology to be answered. Formalizations of human anatomy Adult human anatomy has been catalogued in two very different ways. This very rich resource includes, within a single graph, 94,000 classes and over 170,000 terms, integrated to give approximately 2. Whilst anatomical terminology relating to the human has changed little in the last century, anatomical concepts have evolved much more rapidly and there is great value in novel descriptions and views. For example, the Human Phenotype Ontology grew by 600 classes between 2009 and 2010. Abnormal anatomy and dysmorphology the accurate description of abnormal anatomy is much more difficult than that of canonical anatomy, or even what might be accepted as normal variation within the population. Some abnormalities are so significantly distinct from canonical anatomy that new terms have traditionally been coined to capture the abnormal qualities of the structure, such as turricephaly or brachytelephalangy. Although such terms have definitions that are obvious to humans, articulating the differences between the normal and the abnormal phenotype in ways that are comprehensive computationally turns out to be difficult, and this leads to problems of relating different abnormalities to each other and to canonical structures. What follows is a brief summary of some of the informatics resources associated with abnormal human anatomy that are currently being developed. Such is the nature of the field that some will be expanded at the expense of others, and future interested readers will need to explore the sites to see how the subject is developing. The history and use of clinical dysmorphology terms are complex and very much dependent on the tradition of training and practice in which the practitioner is raised. This requires any resource aiming to use these terms to capture all synonyms and equivalence classes within e34 the terminology, if it is to be of general use. While an ambitious initiative to standardize descriptions of dysmorphology has begun in the Elements of Morphology project, the most widely used approach currently is the collection and relation of synonyms. There is also the broader context to be considered; computational relations of different human phenotype concepts to each other and to those of other species are important in comparing the effects of genetic variation within and between species, and in linking phenotype variation to gene expression. This integration task will now become increasingly important, if the burgeoning data on human disease and model organism phenotypes are to be used to the greatest advantage. The hierarchies showing lineage and parts are on the left and the relationships included in the ontology are on the right. Using this approach, it is possible, for example, to assert that there is a relationship between clinodactyly in humans, bent digits in the mouse and, more distantly, pectoral fin abnormalities in zebrafish. Recent advances that use model organism data to prioritize candidate genes for rare diseases would not be possible without the formalization and integration of anatomical ontologies (Hoehndorf et al 2013). Over the coming few years, we can expect a start to be made in annotating human data on gene expression and abnormal (diseased) phenotypes to defined elements of anatomy (Schofield and Hancock 2012). Once this has happened, collating and integrating a wide range of human tissue-associated data computationally will become possible (Yue et al 2005).
Subthalamic efferents mediate excessive excitatory drive to the internal globus pallidus and substantia nigra pars reticularis women's health clinic north vancouver buy anastrozole 1 mg fast delivery. Overactivity of basal ganglia output then inhibits the motor thalamus and its excitatory thalamocortical connections menopause the musical lyrics purchase cheap anastrozole on-line. Whilst these usually provide good symptomatic relief for many years women's health questions pregnancy symptoms cheap 1 mg anastrozole fast delivery, eventually they lead to the development of side-effects menopause in men symptoms anastrozole 1 mg order without prescription, including dyskinesias menopause symptoms after hysterectomy generic 1 mg anastrozole free shipping. This leads to physiological inhibition of the subthalamic nucleus by overactive pallidosubthalamic neurones. Underactivity of the subthalamic nucleus removes the excitatory drive from internal pallidal neurones, which are known to be underactive in dyskinesias (Crossman 1990). Once again, this anatomical model of basal ganglia function is an oversimplification. Whilst it is true that underactivity of the internal globus pallidus is associated with dyskinesias, it is also known that lesions of the globus pallidus alleviate them. There is evidence that dysfunction of the basal ganglia is also involved in other complex, less well understood, behavioural disorders. In animal experiments, lesions of the basal ganglia, especially of the caudate nucleus, induce uncontrollable hyperactivity. The globus pallidus and thalamus were favoured targets for chemical or thermal lesions. Pallidotomy and thalamotomy often improved rigidity and tremor, but they produced little consistent beneficial effect on akinesia. These lesions were found to be extremely reliable in abolishing contralateral rigidity and drug-induced dyskinesias, with slightly less efficacy on tremor and bradykinesia (Laitinen et al 1992). Implantation of deep-brain electrodes, through which highfrequency pulses generated by a pacemaker could inhibit cells in the vicinity, has been a concept since the early 1970s but did not become a widespread reality until the late 1980s, as a result of technological advances. Subthalamic nucleus stimulation is favoured by most for relieving akinesia and rigidity, with pallidal stimulation considered to ameliorate dyskinesias. Subthalamic stimulation is more effective than pallidal stimulation in allowing patients to reduce their anti-parkinsonian medication. Tremor is best relieved by stimulation of either the contralateral ventral intermediate thalamic nucleus or the zona incerta. Another manifestation of basal ganglia dysfunction is dystonia, which is characterized by increased muscle tone and abnormal postures. Although the pathophysiological basis of dystonia is unclear (Hallett 2006), it is probably caused by underactivity of basal ganglia output. The observation that painful dystonic posturing of the limbs in parkinsonian patients responds dramatically to bilateral pallidal stimulation led to the development of bilateral pallidal stimulation for dystonia. Intriguingly, in dystonia the pallidal neurones are held to fire at rates below normal, and so it is open to question how this stimulation works. Low-frequency stimulation alleviates postural instability and on-medication gait freezing and falling, symptoms that conventional medication and surgery fail to improve. The relative extent of the pedunculopontine nucleus has been outlined based on choline acetyltransferase immunohistochemistry. An explanation of the abnormalities in basal ganglia function involved in dystonia. A review presenting a persuasive argument for functional similarities and intimate reciprocal connections between pedunculopontine nucleus and other basal ganglia structures. A landmark publication setting out a conceptual framework for the way in which the basal ganglia and cerebral cortex process different types of information through largely distinct parallel circuits based on known anatomical connectivity. A review that highlights some limitations of the anatomical model of basal ganglia function. They contain the primary motor and sensory cortices, the highest levels at which motor activities are controlled and to which general and special sensory systems project, and which provide the neural substrate for the conscious experience of sensory stimuli. Association areas are both modality-specific and multimodal, enabling complex analyses of the internal and external environment and of the relationship of the individual with the external world. The elements of the limbic system are particularly concerned with memory and the emotional aspects of behaviour, and provide an affective overtone to conscious experience as well as an interface with subcortical areas such as the hypothalamus, through which widespread physiological activities are integrated. Other cortical areas, primarily within the frontal region, are concerned with the highest aspects of cognitive function and contribute to personality, judgment, foresight and planning. The configuration of the main cerebral sulci and gyri provides the basis for dividing the hemispheres into frontal, parietal, occipital, temporal, insular and limbic lobes. The internal white matter contains association fibres limited to each hemisphere, commissural fibres linking corresponding areas of both hemispheres, and projection fibres connecting the cerebral cortex of each hemisphere with subcortical, brainstem and spinal cord nuclei. Some of these bundles (tracts, fasciculi) are relatively well defined macroscopically and microscopically, while others are less easy to identify. A detailed knowledge of the threedimensional anatomical interrelationships of white matter tracts is a requisite for the planning, intraoperative monitoring and execution of neurosurgical resective procedures. Current understanding of these relationships owes much to the seminal work of Josef Klingler and his meticulous dissection of white matter tracts using formalinfixed, freeze-thawed brains (Agrawal et al 2011). The superolateral surface is convex and lies beneath the bones of the cranial vault; the frontal, parietal, temporal and occipital lobes correspond approximately in surface extent to the overlying bones from which they take their names. The frontal and parietal lobes are separated from the temporal lobe by the prominent lateral (Sylvian) fissure. The inferior surface is divided by the anterior part of the lateral fissure into a small anterior orbital part and a larger posterior tentorial part. The orbital part is the concave orbital surface of the frontal lobe and rests on the floor of the anterior cranial fossa. The posterior part is formed by the basal aspects of the temporal and occipital lobes, and rests on the floor of the middle cranial fossa and the upper surface of the tentorium cerebelli, which separates it from the superior surface of the cerebellum. The medial surface is flat and vertical, separated from the opposite hemisphere by the longitudinal fissure and the falx cerebri. Anteriorly, the cerebral hemisphere terminates at the frontal and temporal poles, and posteriorly at the occipital pole. The cerebral sulci delineate the brain gyri and are extensions of the subarachnoid space (Butler and Hodos 2005, Sarnat and Netsky 1981, Park et al 2007, Chi et al 1977, Nishikuni and Ribas 2013, Ono et al 1990, Catani and Thiebaut de Schotten 2012, Duvernoy 1991, Naidich et al 2013). The main sulci have depths of 13 cm, and their walls harbour small gyri that connect with each other (transverse gyri). Sulci that separate the transverse gyri vary in length and depth, and may become visible as incisures at the surface of the brain. The sulci of the superolateral and inferior surfaces of the hemisphere are usually orientated towards the nearest ventricular cavity. Sulci that are usually continuous include the lateral fissure and the callosal, calcarine, parieto-occipital, collateral and, generally, the central sulcus. On the superolateral surface of the hemisphere, the frontal and temporal regions are each composed of three horizontal gyri (superior, middle and inferior frontal and temporal gyri). The central area is composed of two slightly oblique gyri (pre- and postcentral gyri). The occipital region is composed of two or three less well-defined gyri (superior, middle and inferior occipital gyri). The orbital part of the inferior surface is covered by the orbital gyri and the basal aspect of the rectus gyri, and the tentorial part of the inferior surface is covered by the basal aspects of the inferior temporal, inferior occipital and lingual gyri, and the fusiform gyrus. Non-pyramidal cells, also called stellate or granule cells, are divided into spiny and non-spiny types. The extent to which this organization aids the understanding of cortical functional organization is debatable, but the use of cytoarchitectonic description to identify regions of cortex is common. These are the molecular or plexiform layer; external granular lamina; external pyramidal lamina; internal granular lamina; internal pyramidal (ganglionic) lamina; and multiform (or fusiform/pleiomorphic) layer. Homotypical variants, in which all six laminae are found, are called frontal, parietal and polar, names that link them with specific cortical regions in a somewhat misleading manner. Large pyramidal neurones are found in the greatest densities in agranular cortex, which is typified by the numerous efferent projections of pyramidal cell axons. Although it is often equated with motor cortical areas such as the precentral gyrus (area 4), agranular cortex also occurs elsewhere. In the granular type of cortex the granular layers are maximally developed and contain densely packed stellate cells, among which small pyramidal neurones are dispersed. However, it does receive efferent fibres, derived from the scattered pyramidal cells, although they are less numerous than elsewhere. Granular cortex occurs in the postcentral gyrus (somatosensory area), striate area (visual area) and superior temporal gyrus (acoustic area), and in small areas of the parahippocampal gyrus. Despite its very high density of stellate cells, especially in the striate area, it is almost the thinnest of the five main types. The relative prominence of these major forms of neurone varies reciprocally wherever this form of cortex exists. The parietal type of cortex contains pyramidal cells, which are mostly smaller in size than in the frontal type. In marked contrast, the granular laminae are wider and contain more of the stellate cells: this kind of cortex occupies large areas in the parietal and temporal lobes. The polar type is classically identified with small areas near the frontal and occipital poles, and is the thinnest form of cortex. This is most clearly seen in the lamination of cortical efferent (pyramidal) cells. The internal pyramidal lamina, layer V, gives rise to cortical projection fibres, most notably corticostriate, corticobulbar (including corticopontine) and corticospinal axons. While all are said to develop from the same six-layered pattern, Cerebral hemispheres Four main types of sulci have been described: large primary sulci. Sulci often have side branches that may be unconnected or connected (with end-to-side, end-to-end or side-to-side connections that can also join two neighbouring parallel sulci). Pyramidal cells have a flask-shaped or triangular cell body ranging from 10 to 80 µm in diameter. The soma gives rise to a single thick apical dendrite and multiple basal dendrites. The apical dendrite ascends towards the cortical surface, tapering and branching, to end in a spray of terminal twigs in the most superficial lamina, the molecular layer. From the basal surface of the cell body, dendrites spread more horizontally, for distances up to 1 mm for the largest pyramidal cells. Like the apical dendrite, the basal dendrites branch profusely along their length. A single slender axon arises from the axon hillock, which is usually situated centrally on the basal surface of the pyramidal neurone. Ultimately, in the vast majority of cases, if not in all, the axon leaves the cortical grey matter to enter the white matter. They use an excitatory amino acid, either glutamate or aspartate, as their neurotransmitter. They have relatively small multipolar cell bodies, commonly 6 to 10 µm in diameter. Several primary dendrites, profusely covered in spines, radiate for variable distances from the cell body. The smallest group comprises the heterogeneous non-spiny or sparsely spinous stellate cells. In morphological terms, this is not a single class of cell but a multitude of different forms, including basket, chandelier, double bouquet, neurogliaform, bipolar/fusiform and horizontal cells. Neurones with mainly horizontally dispersed axons include basket and horizontal cells. Basket cells have a short, vertical axon, which rapidly divides into horizontal collaterals, and these end in large terminal sprays synapsing with the somata and proximal dendrites of pyramidal cells. They are small and fusiform, and their dendrites spread short distances in two opposite directions in lamina I. Their axons often stem from a dendrite, then divide into two branches, which travel away from each other for great distances in the same layer. Neurones with an axonal arborization predominantly perpendicular to the pial surface include chandelier, double bouquet and bipolar/ fusiform cells. Chandelier cells have a variable morphology, although most are ovoid or fusiform and their dendrites arise from the upper and lower poles of the cell body. The axonal arborization, which emerges from the cell body or a proximal dendrite, is characteristic and identifies these neurones. The axons ramify close to the parent cell body and terminate in numerous vertically orientated strings, which run alongside the axon hillocks of pyramidal cells, with which they synapse. Generally, these neurones have two or three main dendrites, which give rise to a superficial and deep dendritic tuft. A single axon arises usually from the oval or spindle-shaped cell soma and rapidly divides into an ascending and descending branch. These branches collateralize extensively, but the axonal arbor is confined to a perpendicularly extended, but horizontally confined, cylinder, 5080 µm across. Bipolar cells are ovoid with a single ascending and a single descending dendrite, which arise from the upper and lower poles, respectively. These primary dendrites branch sparsely and their branches run vertically to produce a narrow dendritic tree, rarely more than 10 µm across, which may extend through most of the cortical thickness. Commonly, the axon originates from one of the primary dendrites, and rapidly branches to give a vertically elongated, horizontally confined axonal arbor, which closely parallels the dendritic tree in extent. The frontal and temporal regions each consist of three horizontal gyri; the central area consists of two slightly oblique gyri; the parietal region consists of two lobules (a quadrangular superior lobule and an inferior lobule consisting of two semicircular gyri); the occipital region consists of three irregular, less well defined, predominantly longitudinal gyri that converge towards the occipital pole; and the insula is composed of four or five diagonal gyri. Medially, the external lateral gyri and lobules extend along the superior and inferolateral borders of each hemisphere. Together, they constitute an outer medial ring that surrounds a well-defined, C-shaped inner ring composed of two continuous gyri. Seven to ten thin dendrites typically radiate out from the cell soma, some branching once or twice to form a spherical dendritic field of approximately 100150 µm diameter. Almost immediately, it branches profusely within the vicinity of the dendritic field (and usually somewhat beyond), to give a spherical axonal arbor up to 350 µm in diameter. This is almost certainly the case for basket, chandelier, double bouquet, neurogliaform and bipolar cells. These are afferent fibres, which arise from outside the cortical area, together with intrinsic fibres from cortical interneurones, and the apical dendritic arbors of virtually all pyramidal neurones of the cerebral cortex.
Neural crest cells are induced to differentiate into sympathetic neurones by the dorsal aorta through the actions of the signalling molecules Bmp-4 and Bmp-7 pregnancy glow discount anastrozole 1 mg line. The sympathetic trunk can be seen in the lower cervical and thoracic regions by 45 weeks menopause lose weight buy generic anastrozole from india, and the sympathetic outflow from the lumbosacral sympathetic trunk to the sciatic nerve between 18 and 21 weeks (Ashwell and Waite 2012) breast cancer clothing generic anastrozole 1 mg without prescription. There is cell-specific recognition of postganglionic neurones and the growth cones of sympathetic preganglionic neurones menopause quality of life buy genuine anastrozole online. They meet during their growth women's health center fredericksburg va purchase anastrozole 1 mg without prescription, and this may be important in guidance to their appropriate target. The position of postganglionic neurones, and the exit point from the spinal cord of preganglionic neurones, may influence the types of synaptic connections made, and the affinity for particular postganglionic neurones. When a postganglionic neuroblast is in place, it extends axons and dendrites, and synaptogenesis occurs. The earliest axonal outgrowths from the superior cervical ganglion occur at about stage 14; although the axon is the first cell process to appear, the position of the neurones does not apparently influence the appearance of the cell processes. The local environment plays a major role in controlling the appropriate differentiation of the presumptive autonomic ganglion neurones. The identity of the factors responsible for subsequent adrenergic, cholinergic or peptidergic phenotype has yet to be elucidated; it has been proposed that fibronectin and basal lamina components initiate adrenergic phenotypic expression at the expense of melanocyte numbers. Cholinergic characteristics are acquired relatively early and the appropriate phenotypic expression may be promoted by cholinergic differentiation factor and ciliary neurotrophic factor. Neuropeptides are expressed by autonomic neurones in vitro and may be stimulated by various target tissue factors in sympathetic and parasympathetic neurones. Some neuropeptides are expressed more intensely during early stages of ganglion formation. Premigratory neural crest cells are not pre-patterned for specific axial levels; rather, they attain their axial value as they leave the neuraxis. Once within the gut wall, there is a regionally specific pattern of enteric ganglia formation, which may be controlled by the local splanchnopleuric mesenchyme. Cranial neural crest from somite levels 17 contributes to the enteric nervous system, forming both neuroblasts and glial support cells. The most caudal derivatives of neural crest cells, from the lumbosacral region, somites 28 onwards, form components of the pelvic plexus after migrating through the somites towards the level of the colon, rectum and cloaca. Initially, the cells come to lie within the developing mesentery, then transiently between the layers of the differentiating muscularis externa, before finally forming a more substantial intramural plexus characteristic of the adult enteric nervous system. The gut has a partially established enteric nervous system with a full complement of enteric neurotransmitters by 24 weeks. The rectum is always affected and the proximal extent of the condition is variable. The condition is also characterized by a transition zone with reduced innervation. It is important to be aware that the transition zone may follow an asymmetric course around the circumference of the bowel and a single biopsy may be unreliable. Of the neural crest cells that colonize the bowel, some in the foregut may acquire the ability to migrate outwards and colonize the developing pancreas. Chromaffin cells Chromaffin cells are derived from the neural crest and found at numerous sites throughout the body. They are the classic chromaffin cells of the suprarenal medulla, bronchial neuroepithelial cells, dispersed epithelial endocrine cells of the gut (formerly known as argentaffin cells), carotid body cells, and the paraganglia (Ch. The sympathetic ganglia, suprarenal medulla and chromaffin cells are all derived from the cells of the sympathosuprarenal lineage. In the suprarenal medulla, these cells differentiate into a number of types consisting of small and intermediate-sized neuroblasts or sympathoblasts, and larger, initially rounded phaeochromocytoblasts. Large cells with pale nuclei, thought to be the progenitors of chromaffin cells, can be detected from 9 weeks in human fetuses, and clusters of small neurones are evident from 14 weeks. Intermediate-sized neuroblasts differentiate into the typical multipolar postganglionic sympathetic neurones (which secrete noradrenaline (norepinephrine) at their terminals) of classic autonomic neuroanatomy. The large cells differentiate into masses of columnar or polyhedral phaeochromocytes (classic chromaffin cells), which secrete either adrenaline (epinephrine) or noradrenaline (norepinephrine). These cell masses are termed paraganglia and may be situated near, on the surface of, or embedded in, the capsules of the ganglia of the sympathetic chain, or in some of the large autonomic plexuses. The largest members of the latter are the para-aortic bodies that lie along the sides of the abdominal aorta in relation to the inferior mesenteric artery. During childhood, the para-aortic bodies and the paraganglia of the sympathetic chain partly degenerate and can no longer be isolated by gross dissection, but even in the adult, chromaffin tissue can still be recognized microscopically in these various sites. Enteric nervous system the enteric nervous system is different from the other components of the autonomic nervous system because it can mediate reflex activity independently of control by the brain and spinal cord. The neuroblasts of the lateral walls of the tube are large and, at first, round or oval (apolar). However, one process is withdrawn and the neuroblast becomes unipolar, although this is not invariably so in the case of the spinal cord. Further differentiation leads to the development of dendritic processes and the cells become typical multipolar neurones. In the developing cord, they occur in small clusters representing clones of neurones. Caudally, the central canal of the cord ends as a fusiform dilation, the terminal ventricle. In the human embryo, the definitive grouping of the ventral column cells, which characterizes the mature cord, occurs early; by the fourteenth week (80 mm), all the major groups can be recognized. As the anterior and lateral grey columns assume their final form, the germinal cells in the ventral part of the ventricular zone gradually stop dividing. The layer becomes reduced in thickness until, ultimately, it forms the single-layered ependyma that lines the ventral part of the central canal of the spinal cord. In this way, the central canal becomes relatively reduced in size and somewhat triangular in outline. At about the end of the fourth week, advancing axonal sprouts invade the marginal zone. The first to develop are those destined to become short intersegmental fibres from the neuroblasts in the intermediate zone, and fibres of dorsal roots of spinal nerves that pass into the spinal cord from neuroblasts of the early spinal ganglia. The earlier dorsal root fibres that invade the dorsal marginal zone arise from small dorsal root ganglionic neuroblasts. This bundle increases in size and, spreading towards the median plane, forms the primitive posterior funiculus of fine calibre. Later, fibres derived from new populations of large dorsal root ganglionic neuroblasts join the dorsal root; they are destined to become fibres of much larger calibre. As the posterior funiculi increase in thickness, their medial surfaces come into contact, separated only by the posterior medial septum, which is ependymal in origin and neuroglial in nature. It is thought that the displaced primitive posterior funiculus may form the basis of the dorsolateral tract or fasciculus (of Lissauer). Their disposition may be determined in part by contact guidance along the earliest radial array of glial fibres that cross the full thickness of the early neuroepithelium. This enlargement outlines the anterior (ventral) column of the grey matter and causes a ventral projection on each side of the median plane; the floor plate remains at the bottom of the shallow groove so produced. As growth proceeds, these enlargements, which are further increased by the development of the anterior funiculi (tracts of axons passing to and from the brain), encroach on the groove until it becomes converted into the slit-like anterior median fissure of the adult spinal cord. The axons of some of the neuroblasts in the anterior grey column cross the marginal zone and emerge as bundles of ventral spinal nerve rootlets on the anterolateral aspect of the spinal cord. These constitute, eventually, both the -efferents that establish motor end-plates on extrafusal striated muscle fibres and the -efferents that innervate the contractile polar regions of the intrafusal muscle fibres of the muscle spindles. Maturation of the spinal cord Long intersegmental fibres begin to appear at about the third month and corticospinal fibres are seen at about the fifth month. In many sites, slow growth continues for long periods, even into the postpubertal years. The cervical and lumbar enlargements appear at the time of the development of their respective limb buds. Lateral grey column In the thoracic and upper lumbar regions, some intermediate zone neuroblasts in the dorsal part of the basal plate outline a lateral column. Their axons join the emerging ventral nerve roots and pass as preganglionic fibres to the ganglia of the sympathetic trunk or related ganglia, the majority eventually myelinating to form white rami communicantes. The axons within the rami synapse on the autonomic ganglionic neurones, and axons of some of the latter pass as postganglionic fibres to innervate smooth muscle cells, adipose tissue or glandular cells. Other preganglionic sympathetic efferent axons pass to the cells of the suprarenal medulla. It gives origin to the preganglionic parasympathetic fibres that run in the pelvic splanchnic nerves. The anterior region of each basal plate initially forms a continuous column of cells throughout the length of the developing cord. This soon develops into two columns (on each side): one is medially placed and concerned with innervation of axial musculature, and the other is laterally placed and innervates the limbs. At limb levels, the lateral column enlarges enormously, but regresses at other levels. Numerous interneurones develop in these sites (including Renshaw cells); it is uncertain how many of these differentiate directly from ventrolateral lamina (basal plate) neuroblasts and how Ascent of conus medullaris 252 In early embryonic life, the spinal cord occupies the entire length of the vertebral canal, and the spinal nerves pass at right angles to the cord. From the second trimester, the vertebral column grows more rapidly than the spinal cord; the relationship between the conus medullaris and the vertebral column therefore changes as the conus gradually ascends to lie at higher vertebral levels (Barson 1970). Most of this relative rostral migration occurs during the first half of intrauterine life. By the twenty-fifth week, the terminal ventricle of the spinal cord has altered in level from the second coccygeal vertebra to the third lumbar, a distance of nine segments. As the change in level begins rostrally, the caudal end of the terminal ventricle, which is adherent to the overlying ectoderm, remains in situ, and the walls of the intermediate part of the ventricle and its covering pia mater become drawn out to form a delicate filament, the filum terminale. The separated portion of the terminal ventricle persists for a time, but it usually disappears before birth. It does, however, occasionally give rise to congenital cysts in the neighbourhood of the coccyx. In the definitive state, the upper cervical spinal nerves retain their position roughly at right angles to the cord. Proceeding caudally, the nerve roots lengthen and become progressively more oblique. By full term (40 weeks), the conus medullaris lies between the first and third lumbar vertebrae. Central nervous system Determining the prenatal position of the conus medullaris relative to specific vertebrae is a widely used method of identifying the termination of the spinal cord. The often wide discrepancies between published values attest to the fact that ageing preterm infants remains an inexact science. The differentiation of the lateral walls of the hindbrain into basal (ventrolateral) and alar (dorsolateral) plates has a similar significance to the corresponding differentiation in the lateral wall of the spinal cord, and ventricular, intermediate and marginal zones are formed in the same way. Cells of the basal plate (ventrolateral lamina) Rhombencephalon By the time the midbrain flexure appears, the length of the rhombencephalon is greater than that of the combined extent of the mesencephalon and prosencephalon. Ventrally, the hindbrain is separated from the dorsal wall of the primitive pharynx only by the notochord, the two dorsal aortae and a small amount of mesenchyme; on each side, it is closely related to the dorsal ends of the pharyngeal arches. The greatest increase in width corresponds to the region of maximum convexity, so that the outline of the roof plate becomes rhomboidal. By the same change, the lateral walls become separated, particularly dorsally, and the cavity of the hindbrain, subsequently the fourth ventricle, becomes flattened and somewhat triangular in cross-section. At about 4 1 2 weeks of development, when the pontine flexure is first discernible, the association between the rhombomeres and the underlying motor nuclei of certain cranial nerves can be seen. The general pattern of distribution of motor nuclei is as follows: rhombomere 1 contains the trochlear nucleus, rhombomeres 2 and 3 contain the trigeminal nucleus, rhombomeres 4 and 5 contain the facial nucleus, rhombomere 5 contains the abducens nucleus, rhombomeres 6 and 7 contain the glossopharyngeal nucleus, and rhombomeres 7 and 8 contain the vagal, accessory and hypoglossal nuclei. The general visceral afferent column is represented by a part of the dorsal nucleus of the vagus, the special visceral afferent column by the nucleus of the tractus solitarius, the general somatic afferent column by the afferent nuclei of the trigeminal nerve, and the special somatic afferent column by the nuclei of the vestibulocochlear nerve. Postganglionic neurones are associated with the general visceral efferent column, bipolar neurones are associated with the otocyst, and unipolar afferent neurones are associated with the other alar plate columns. It is represented in the caudal part of the hindbrain by the hypoglossal nucleus, and it reappears at a higher level as the nuclei of the abducens, trochlear and oculomotor nerves (somatic efferent nuclei). The intermediate column is represented in the upper part of the spinal cord and caudal brainstem (medulla oblongata and pons), and its neurones supply branchial (pharyngeal) and postbranchial musculature. It is discontinuous, forming the elongated nucleus ambiguus in the caudal brainstem, which gives fibres to the ninth, tenth and eleventh cranial nerves, and continues into the cervical spinal cord as the origin of the accessory nerve. At higher levels, parts of this column give origin to the motor nuclei of the facial and trigeminal nerves. The nucleus ambiguus and the facial and trigeminal motor nuclei are termed branchial (special visceral) efferent nuclei. Neurones in the most dorsal column of the basal plate (represented in the spinal cord by the lateral grey column) innervate viscera. The column is discontinuous; its large caudal part forms some of the dorsal nucleus of the vagus and its cranial part forms the salivatory nucleus. These nuclei are termed general visceral (general splanchnic) efferent nuclei, and their neurones give rise to preganglionic, parasympathetic nerve fibres. It is important to note that the neurones of the basal plate and their three columnar derivatives are only motor in the sense that some of their number form either motor neurones or preganglionic parasympathetic neurones. The remainder, which greatly outnumber the former, differentiate into functionally related interneurones and, in some loci, into neuroendocrine cells. The emergent neurobiological mechanisms are, in fact, much more complex and less well understood. It has been suggested that a neurone tends to remain as near as possible to its predominant source of stimulation, and that, to achieve this aim, it will migrate around intervening structures, towards the greatest density of stimuli. The curious paths of the axons arising from the facial nucleus and the nucleus ambiguus have been regarded as exemplars of this phenomenon of neurobiotaxis. In a 10 mm embryo, the facial nucleus lies in the floor of the fourth ventricle, occupying the position of the special visceral efferent column, and it is placed at a higher level than the abducens nucleus. As growth proceeds, the facial nucleus migrates at first caudally and dorsally, relative to the abducens nucleus, and then ventrally to reach its adult position. As it migrates, the axons to which its somata give rise elongate and their subsequent course is assumed to map out the pathway along which the facial nucleus has travelled.
Generic 1 mg anastrozole overnight delivery. MY VAGINA MASSAGE!.
References
- Francis WJA, Jeffcoate TNA. Dyspareunia following vaginal operations. Br J Obstet Gynaecol 1961; 68: 1.
- Vital C, Vital A, Dupon M, Gin H, Rouanet-Larriviere M, Lacut JY. Acute painful diabetic neuropathy: Two patients with recent insulin-dependent diabetes mellitus. J Peripher Nerv Syst. 1997;2:151-154.
- Neary DF, Snowden JS, Gustafson L, et al. Frontotemporal lobar degeneration: a consensus on clinical diagnostic criteria. Neurology. 1998;51:1546-1554.
- Colloca L, Benedetti F. Placebo analgesia induced by social observational learning. Pain. 2009;144:28-34.
- Calin A. Terminology, introduction, diagnostic criteria and overview. In: The Spondylarthritides. Eds Calin A and Taurog JD. OUP, Oxford 1998; pp3-6. Khan MA, van der Linden SM. A wider spectrum of spondylarthropathies. Sem Arthritis Rheum 1990; 20: 107n13.
- Lenhardt R, Marker A, Goli V et al. Mild intra-oerative hyothermia rolongs ostanaesthetic recovery. Anaesthesiology 1997;87:1318-23 31.