

Sumycin
| Contato
Página Inicial
Deirdre Pallister
- Associate specialist breast clinician
- Jarvis Breast Screening Centre, Guildford
- and Royal Marsden Hospital, London, UK
In vitro antibacterial activity of tigecycline in comparison with doxycycline antimicrobial drugs 250 mg sumycin purchase free shipping, ciprofloxacin and rifampicin against Brucella spp antibiotics used for cellulitis order sumycin amex. In vitro activities of tigecycline and 10 other antimicrobials against nonpigmented rapidly growing mycobacteria antibiotics in copd exacerbation discount sumycin master card. Safety and efficacy of tigecycline in treatment of skin and skin structure infections: results of a double-blind phase 3 comparison study with vancomycin-aztreonam antibiotics for sinus infection in horses 250 mg sumycin with amex. Efficacy and safety of tigecycline monotherapy compared with vancomycin plus aztreonam in patients with complicated skin and skin structure infections: results from a phase 3 antimicrobial 2008 sumycin 500 mg purchase without a prescription, randomized, double-blind trial. Overview of tigecycline efficacy and safety in the treatment of complicated skin and skin structure infections-a European perspective. Efficacy and safety of tigecycline monotherapy compared with vancomycinaztreonam in the treatment of complicated skin and skin structure infections in patients from India and Taiwan. A multicentre, open-label, randomized comparative study of tigecycline versus ceftriaxone sodium plus metronidazole for the treatment of hospitalized subjects with complicated intraabdominal infections. Efficacy of tigecycline versus ceftriaxone plus metronidazole for the treatment of complicated intra-abdominal infections: results from a randomized, controlled trial. Efficacy and safety of tigecycline versus levofloxacin for communityacquired pneumonia. Safety and efficacy of intravenous tigecycline in treatment of communityacquired pneumonia: results from a double-blind randomized phase 3 comparison study with levofloxacin. Tigecycline in the treatment of ventilator-associated pneumonia: experience from the Latin American Tigecycline Use Registry. Comparison of tigecycline with imipenem/cilastatin for the treatment of hospital-acquired pneumonia. Indications of a new antibiotic in clinical practice: results of the tigecycline initial use registry. Pilot trial of N-acetylcysteine and tigecycline as a catheter-lock solution for treatment of hemodialysis catheter-associated bacteremia. A Phase 3, openlabel, non-comparative study of tigecycline in the treatment of patients with selected serious infections due to resistant gram-negative organisms including Enterobacter species, Acinetobacter baumannii and Klebsiella pneu moniae. Efficacy and safety of tigecycline compared with vancomycin or linezolid for treatment of serious infections with methicillin-resistant Staphylococcus aureus or vancomycin-resistant enterococci: a phase 3, multicentre, double-blind, randomized study. Tigecycline use in cancer patients with serious infections: a report on 110 cases from a single institution. Comparative effectiveness of aminoglycosides, polymyxin B, and tigecycline for clearance of carbapenem-resistant Klebsiella pneu moniae from urine. A novel glycylcycline, 9-(N,N-dimethylglyc ylamido)-6-demethyl-6deoxytetracycline, is neither transported nor recognized by the transposon Tn10-encoded metal-tetracycline/H+ antiporter. Tigecycline is efficacious in the treatment of complicated intra-abdominal infections. Results of a multicenter, randomized, open-label efficacy and safety study of two doses of tigecycline for complicated skin and skinstructure infections in hospitalized patients. Tigecyclineinduced acute pancreatitis: about two cases and review of the literature. Systematic review and metaanalysis of the effectiveness and safety of tigecycline for treatment of infectious disease. Efficacy and safety of tigecycline for the treatment of infectious diseases: a meta-analysis. A fatal case of aplastic anemia following chloramphenicol (chloromycetin) therapy. Recovery of polysome function of T4-infected Escherichia coli after brief treatment with chloramphenicol and rifampin. Bactericidal and bacteriostatic action of chloramphenicol against memingeal pathogens. Chloramphenicolinduced erythroid suppression and bone marrow ferrochelatase activity in dogs. Pharmacokinetics of chloramphenicol and chloramphenicol succinate in infants and children. Pharmacokinetic comparison of intravenous and oral chloramphenicol in patients with Haemophilus influenzae meningitis. Removal and absorption of antibiotics in patients with renal failure undergoing peritoneal dialysis. Evaluation of chloramphenicol acid succinate therapy of induced typhoid fever and Rocky Mountain spotted fever. Chloramphenicol and its metabolic products in the blood of patients with severe renal disease or hepatic cirrhosis. Antibiotic concentrations in ascitic fluid of patients with ascites and bacterial peritonitis. The permeability of traumatically inflamed synovial membrane to commonly used antibiotics. In vitro activity of thiamphenicol against multiresistant Streptococcus pneu moniae, Haemophilus influenzae and Staphylococcus aureus in Italy. High-level vancomycin-resistant Staphylococcus aureus isolates associated with a polymicrobial biofilm. Susceptibility of Nocardia asteroides to 46 antibiotics, including 22 betalactams. In vitro activities of ciprofloxacin, cefotaxime, ceftriaxone, chloramphenicol, and rifampin against fully susceptible and moderately penicillin-resistant Neisseria meningitidis. Drug susceptibility of Neisseria isolates from patients attending clinics for sexually transmitted diseases in Addis Ababa. Antibiotic resistance of faecal Escherichia coli from healthy volunteers from eight developing countries. Resistance among Escherichia coli to sulphonamides and other antimicrobials now little used in man. Susceptibility of "enterobacteria" to aminoglycoside antibiotics: comparisons with tetracyclines, polymyxins, chloramphenicol, and spectinomycin. In vitro activity of cinoxacin, ampicillin, and chloramphenicol against Shigella and nontyphoid Salmonella. Antimicrobial susceptibility of respiratory isolates of Enterobacteriaceae and Staphylococcus aureus in Italy: incidence and trends over the period 1997-1999. Antimicrobial susceptibility of pathogenic Yersinia enterocolitica isolated in Canada from 1972 to 1990. Role of efflux pump(s) in intrinsic resistance of Pseudomonas aeruginosa: active efflux as a contributing factor to beta-lactam resistance. Antibiotic resistance patterns and extended-spectrum beta-lactamase production among Acinetobacter spp. Chloramphenicol resistance in Pseudomonas cepacia because of decreased permeability. Hybridization analysis of three chloramphenicol resistance determinants from Clostridium perfringens and Clostridium difficile. Correlation between serogroup and susceptibility to chloramphenicol, clindamycin, erythromycin, rifampicin and tetracycline among 308 isolates of Clostridium difficile. National survey on the susceptibility of Bacteroides fragilis group: report and analysis of trends in the United States from 1997 to 2004. Mycoplasma pneu moniae disease: clinical spectrum, pathophysiology, epidemiology, and control. Response to antimicrobial therapy in childhood bacterial meningitis in tropical Africa: report of a bi-centre experience in Nigeria, 1993-1998. Chloramphenicol or ceftriaxone, or both, as treatment for meningitis in developing countries Failure of chloramphenicol therapy in penicillin-resistant pneumococcal meningitis. Multi-antibiotic resistant brain abscess sensitive only to chloramphenicol: a case report. Treatment of typhoid fever in children with a flexible-duration of ceftriaxone, compared with 14-day treatment with chloramphenicol. Comparative efficacy of chloramphenicol, ampicillin, and co-trimoxazole in the treatment of typhoid fever. Effect of antibiotic therapy in acute salmonellosis on the fecal excretion of salmonellae. Analysis of risk factors for fatal Rocky Mountain Spotted Fever: evidence for superiority of tetracyclines for therapy. Chloramphenicol for the treatment of vancomycin-resistant enterococcal infections. Chloramphenicol treatment for vancomycin-resistant Enterococcus faecium bacteremia. Chloramphenicol treatment for acute infective conjunctivitis in children in primary care: a randomised double-blind placebo-controlled trial. Resistance mechanism of chloramphenicol in Streptococcus haemolyticus, Streptococcus pneumoniae and Streptococcus faecalis. Salmonella typhi in vivo acquires resistance to both chloramphenicol and co-trimoxazole. Cloning and expression in Escherichia coli of a gene encoding nonenzymatic chloramphenicol resistance from Pseudo monas aeruginosa. A permeability barrier as a mechanism of chloramphenicol resistance in Haemophilus influenzae. Mobile genes coding for efflux-mediated antimicrobial resistance in gram-positive and gram-negative bacteria. Active efflux of chloramphenicol in susceptible Escherichia coli strains and in multiple-antibiotic-resistant (Mar) mutants. Chloramphenicol-induced hemolysis in Caucasian glucose-6-phosphate dehydrogenase deficiency. Chloramphenicol-induced bone marrow injury: possible role of bacterial metabolites of chloramphenicol. Aplastic anemia associated with parenteral chloramphenicol: review of 10 cases, including the second case of possible increased risk with cimetidine. Risk of serious haematological toxicity with use of chloramphenicol eye drops in a British general practice database. Possible association between ocular chloramphenicol and aplastic anaemia- the absolute risk is very low. Fatal cardiovascular collapse of infants receiving large amounts of chloramphenicol. Acute myocardial effects of chloramphenicol in newborn pigs: a possible insight into the gray baby syndrome. The inhibitory effect of chloramphenicol on the synthesis of antibody in tissue culture. Susceptibility of Esche richia coli K1 to four combinations of antimicrobial agents potentially useful for treatment of neonatal meningitis. Antagonism by chloramphenicol of broad-spectrum beta-lactam antibiotics against Klebsiella pneumoniae. Activity of gatifloxacin and ciprofloxacin in combination with other antimicrobial agents. Nicknamed "Rififi" after a French crime movie, the official name became rifamycin. Rifampin is increasingly utilized as an adjunct for foreign-body infections associated with biofilm production, in combination with other antibiotics for infections due to multidrug resistant gram-negative bacilli, and for infections caused by intracellular pathogens. However, pathogens develop resistance to rifampin at a rate of 10-8 to 10-9 per bacterium per cell division. In particular, amino acid substitutions at positions 526 and 531 conferred high-level resistance to rifampin, rifabutin, and rifapentine. Finally, the enzyme from some species, such as Nocardia, is intrinsically less susceptible to rifampin inhibition because of metabolic changes at the critical hydrogen-binding sites. Gram-negative bacteria against which rifampin is effective include Haemophilus influenzae, Neisseria meningitidis, and Helicobacter pylori. Rifampin is most potent against gram-positive bacteria, including Staphylococcus aureus, coagulase-negative staphylococci, Streptococcus pyogenes, Streptococcus pneumoniae, Streptococcus viridans, C. The drug undergoes rapid and complete absorption, which is improved if taken on an empty stomach. Rifampin undergoes an enterohepatic circulation and deacetylation with rapid elimination in bile. Urinary excretion is 13% to 24% with no dose adjustment necessary in renal insufficiency. The rifampin substrate is red, and the high lipophilic properties and wide distribution often turn body fluids such as urine, tears, sweat, feces, and contact lenses red-orange. Fifty-three percent of the dose is excreted in urine as the primary active C-25 desacetyl metabolite, and dose reduction is recommended for a creatinine clearance less than 30 mL/min. Rifapentine, a cyclopentyl rifamycin, is a more potent and longeracting rifamycin. Rifapentine achieves high intracellular concentrations, exceeding that of rifampin. The primary route of metabolism, mediated by an esterase enzyme, is nonoxidative to the 25-desacetyl metabolite. Rifampin, rifabutin, and rifapentine are metabolized in hepatocytes and intestinal microsomes to deacetylated, hydroxylated, and formyl derivatives. Absorption after an oral dose is minimal secondary to low intestinal permeability and water solubility. Although caution is recommended in using rifaximin in patients with Child-Pugh class C cirrhosis, no dosage adjustments are required. The degree of interaction correlates with the dose and frequency of administration. Clostridium difficile Pseudomonas aeruginosa Mycobacterium tuberculosis Mycobacterium aviumintracellulare Mycobacterium kansasii Mycobacterium marinum Mycobacterium fortuitum Mycobacterium scrofulaceum Legionellaspp. The rifamycins have similar side effect profiles, except for rifabutin, which may cause uveitis and polyarthralgias.
Evolution antibiotics insomnia purchase cheap sumycin, population structure antimicrobial grout sumycin 250 mg low cost, and phylogeography of genetically monomorphic bacterial pathogens virus 3d project 500 mg sumycin visa. A disseminated multidrug-resistant clonal group of uropathogenic Escherichia coli in pyelonephritis virus alert lyrics purchase sumycin 500 mg with amex. Bordetella species are distinguished by patterns of substantial gene loss and host adaptation antibiotic klebsiella order sumycin 500 mg. Intrinsic thermal sensing controls proteolysis of Yersinia virulence regulator RovA. Pathogenic mechanisms of Neisseria gonorrhoeae: observations on damage to human fallopian tubes in organ culture by 67. Surface organelles assembled by secretion systems of gram-negative bacteria: diversity in structure and function. E-cadherin is the receptor for internalin, a surface protein required for entry of L. Translocated intimin receptors (Tir) of Shiga-toxigenic Escherichia coli isolates belonging to serogroups O26, O111, and O157 react with sera from patients with hemolytic-uremic syndrome and exhibit marked sequence heterogeneity. Nuclear factor kappa B protects against host cell apoptosis during Rickettsia rickettsii infection by inhibiting activation of apical and effector caspases and maintaining mitochondrial integrity. Ultravioletinduced cell death blocked by a selenoprotein from a human dermatotropic poxvirus. Signature tagged mutagenesis in the functional genetic analysis of gastrointestinal pathogens. Identifying microbial fitness determinants by insertion sequencing using genome-wide transposon mutant libraries. Recovery of divergent avian bornaviruses from cases of proventricular dilatation disease: identification of a candidate etiologic agent. A culture-independent sequence-based metagenomics approach to the investigation of an outbreak of Shigatoxigenic Escherichia coli O104: H4. A host transcriptional signature for presymptomatic detection of infection in humans exposed to influenza H1N1 or H3N2. Temporal dynamics of the transcriptional response to dengue virus infection in Nicaraguan children. Even if a single microbe is the etiologic agent of infection, the pathogenesis and pathophysiology of infection can be viewed within the context of the microbiome and human biology. We now appreciate that our human microbiome is a complex ecosystem, with distinct biologic niches. The resultant perspective for human health and disease shifts the focus to the global balance of our microbiota rather than the appearance of a specific infectious agent. As a result, a clear understanding of the role of microbial community structure in the host can facilitate a deeper understanding of infectious diseases and susceptibility to infections (see Table 2-1). We are realizing the translational fruits of a broadened understanding of the human microbiome as metagenomic medicine makes strides in restoring health in highly morbid conditions. Several theories have been proposed, such as acquisition from exposure to the maternal vaginal microbiome, intestinal microbiome, breast milk microbiome, and skin-to-skin contact7; however, these findings are not exclusive. In murine models, the newborn gut is relatively sterile before birth and soon after delivery is exposed to both the maternal and environmental microbiota. The mode of delivery has been associated with differences in composition of the newborn microbiome. In one of the often-cited studies regarding mode of delivery and the impact on the human microbiome, investigators examined neonatal microbial community structure in instances of cesarean delivery in comparison with vaginal birth from a small cohort of Venezuelan women employing 16S-based metagenomics. Different body habitats contain microbial communities and microbiomes that differ by microbial composition and function (metabolic modules and pathways). As a result, each body habitat is composed of characteristic bacterial species and other microbial taxa that are adapted to each body site. Differences in microbial composition yield differences in metabolic capacity and aggregate function of the human microbiome. We now appreciate that the microbial genome exceeds the human genome by at least 250-fold, and the cellular count of resident microbiota exceeds the human cell count by greater than 10-fold. Abundance refers to the relative quantity of microbes within each individual or body site, whereas ubiquity refers to the presence of the same microbes in different individuals. Canonical pathogens as defined by the National Institute of Allergy and Infectious Diseases2 are generally absent from the human microbiome in healthy individuals, but opportunistic pathogens are widely distributed in healthy adults. This finding contrasts with the relative habitat specificity of commensal species that lack evidence of pathogenicity. In summary, although canonical pathogens are rare in healthy individuals, opportunistic pathogens are relatively common in healthy individuals and explain why immunosuppression often results in opportunistic infections. Canonical pathogens, by contrast, must be transmitted to healthy individuals from other humans, animals, or the environment. Opportunistic pathogens may arise from within the indigenous microbiome, in addition to possible transmission from outside sources. In this chapter we describe the current state of knowledge of the human microbiome and key features of human-associated microbial communities in each primary body habitat. We render brief discussions regarding known determinants of the microbial structure of these niches and presumptive associations with several disease states (as examples). Because of the paucity of data pertaining to a definitive role of variance in the microbiome as the source of disease, we limit the entirety of our discussion in this chapter to co-occurrence and associations, rather than presumptive causation. Although a significant amount of research has focused on the gut microbiome with respect to health and disease, there is a substantial body of work regarding the oral microbiome. To put this into perspective, 1 cc (mL) of human saliva in a healthy adult contains approximately 100 million cells, which is discrete from the community of the surrounding oral microbiome. Several studies2,3,15-18 have documented the unanticipated robustness of the oral microbiome. Microarray, early pyrosequencing, and culture methodologies estimated approximately 700 oral microbial phylotypes. However, dental plaque sampling pooled from 98 healthy adults was estimated to represent 22 phyla comprising 3621 and 6888 species-level phylotypes in the saliva and plaque, respectively. In terms of the longitudinal establishment of the human microbiome, the microbiomes in vaginally versus cesarean-delivered infants yielded a modest but significant difference at up to 6 months of age and appreciable differences years later. Metagenomic massively parallel sequencing approaches have demonstrated exquisite body site specificity, and higher level. Represented in the figure are relative distributions (percentages) of taxa projected at the phylum level. Although these differences are also observed in the gut and skin (leading to issues such as methicillin-resistant S. The tonsil microbiome can be distinguished from the tongue, and the tongue from the palate. These differences are evident despite spatial proximity and constant contact between these sites. With maintenance of niche and subsite specificity in mind, it is not surprising that long-standing associations have been documented between oral health and disease manifestations in distal body sites. For example, periodontal disease is the most common infectious disease affecting the teeth. Left untreated or ineffectively treated, periodontitis is a known independent predictor of, and comorbid contributor to , preterm birth, cardiovascular disease, pulmonary disorders, diabetes, and obesity. The generation of the dental plaque biofilm that we experience daily has been well characterized. Rather, the complexity of the subgingival microbiota and biofilm establishment promote a model of a microbial community-associated disease. In one recent study using deep sequencing, periodontitis was associated with a shift to populations enriched with gram-negative genera such as Catonella, Haemophilus, and Tannerella. One exception is infection by Aggregatibacter actinomycetemcomitans because this gram-negative rod appears to cause a highly aggressive periodontitis (localized aggressive periodontitis) in Africans with strong host tropism. The human skin comprises various ecosystems that differ markedly by relative differences in temperature, 14 humidity, and glandular distribution. The human skin microbiome and the nature of the local environment can vary greatly depending on anatomic location. One report described bacterial compositional differences in 20 different sites on the human skin. These factors include host physiology (sex, age, site); environment (local climate, geographic location); immune system; host genotype; lifestyle (occupation, hygiene); and pathobiology (skin and systemic diseases). These glands produce oily substances such as sebum and other lipid, carbohydrate, and proteinaceous components that may serve as nutrients for the microbiome, as well as inhibitors to particular classes of microbes. For example, sebaceous glandrich regions include the head, shoulders, upper arms, and upper torso. Apocrine glands are enriched around the eyes and ears, nipples, and genital regions. Relative humidity is another key factor affecting microbial composition of the skin. This study confirmed that the skin microbiome is distinct from that of other body sites and it is characterized by an intermediate degree of alpha diversity and richness per specimen. The phyla Actinobacteria, Firmicutes, and Verrucomicrobia were the dominant groups in the human skin,24 in contrast with the predominance of Bacteroidetes, Firmicutes, and Proteobacteria in the human gut. Therefore, even at the level of phyla composed of hundreds of different bacterial species, stark differences are evident in the skin compared to other body sites. For evaluation of dermatologic infections, deeper surface and tissue sampling may be necessary in order to detect infectious agents. Age is an important factor as evidenced by shifts in bacterial communities that occur during the sexual maturation process. Specific groups of microbes may be conserved in the skin of healthy individuals, whereas interindividual variation may account for differences in relative abundances of microbes and differences in disease susceptibilities. Representative bacterial genera in the human skin across sites include Corynebacterium, Eubacterium, Propionibacterium, Staphylococcus, and Streptococcus,24 as well as the fungal Malassezia spp. The genus Malassezia is the predominant fungal genus of the human skin at multiple body sites, including the head, torso, arms, and legs,24 except for sites on the foot. One study described the utility of skin fingertip microbiome patterns for tracking the use of keyboards and perhaps other devices by specific individuals. The researchers determined that the overall lung community appeared consistent between the various sampling sites but that decreased amounts of bacterial content were isolated from deeper lung specimens. This significant shift in community composition was attributed to an overall increase in the amount of S. Predominant genera were similar across both groups with the most common bacteria identified as Streptococcus, Granulicatella, Actinomyces, Prevotella, and Veillonella. Bacterial and viral pathogens have been implicated as possible causes of asthma and potential triggers of asthmatic episodes. In a study of healthy children and children with asthma, significant shifts in overall bacterial communities present in the respiratory tract were not detected at the phylum level with both groups displaying the expected predominance of Bacteroidetes, Firmicutes, and Proteobacteria (the asthma group exhibited a slightly different order of prevalence: Firmicutes, Proteobacteria, and Bacteroidetes). Adding to the microbiome data in the asthma population is a compelling study from Ecuador, whereby treatment of respiratory illnesses differs greatly from the standard of care in the United States. Oropharyngeal swabs were obtained from both wheezing and healthy infants, and all patients had minimal exposure to antibiotics and no exposure to inhaled steroids. The most common genera isolated were consistent with the findings of Hilty and colleagues,36 with most bacteria identified as Streptococcus, Veillonella, Atopobium, and Prevotella. In the wheezing group, a greater frequency of Neisseria, Prevotella, Corynebacterium, Staphylococcus, Actinomyces, and Haemophilus was detected. Although limited data exist on fungal communities of the airways, Candida, Aspergillus, Geotrichum, and Malassezia sp. Overall, Veillonella and Streptococcus were most commonly identified among both specimen types. Streptococcus, Veillonella, and Prevotella were the most prevalent genera in the respiratory tract, and Bacteroides, Bifidobacterium, and Veillonella were the most prevalent genera in the gastrointestinal tract. In general, bacterial diversity increased over time, with more rapid diversification occurring in the developing respiratory tract. Interestingly, gut colonization preceded subsequent respiratory colonization for several genera, including Roseburia, Dorea, Sporacetigenium, Coprococcus, Blautia, Enterococcus, and Escherichia. Aspiration may account for the spread of organisms from the gut to the airways, possibly resulting in infections in the compromised host. By contrast, the distal esophagus immediately cephalic to the gastroesophageal sphincter contains a moderately diverse microbiome. This esophageal region appears to harbor a collection of permanent residents that include bacteria, yeasts, and viruses in human patients. Older culture-based studies showed that gram-positive bacteria such as Streptococcus dominated the distal esophageal ecosystem. In terms of global parameters, the distal esophageal microbiome is less rich and less diverse than that of the large intestine. The phenotypically normal distal esophagus contains a less complex microbiome composed largely of the phylum Firmicutes and dominated by the genus Streptococcus. To explain these findings, we propose the microbial diversity setpoint hypothesis. This hypothesis states that increased diversity in regions of lesser diversity is associated with disease and inflammation, and reduced diversity in regions of greater diversity is associated with disease and inflammation. Increased bacterial species richness and diversity in the esophagus were associated with esophagitis and Barrett esophagus. Discovery of Helicobacter pylori in 1982 led to the widespread appreciation of bacterial colonization in the human stomach. In summary, these discussions laid the foundation for concepts of the human microbiome and how antibiotics could increase chronic disease risk by extermination of valuable members of the human gastrointestinal microbiome. The dominant phyla included phyla dominant in the large intestine, such as Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria, but the phylum Fusobacteria seems to be differentially enriched in the stomach. An interesting finding was the presence of Deinococcus-like organisms in the stomach, undefined at the time of publication. Differences in gastric bacterial communities may predispose patients to acute or chronic gastritis. The inability to maintain a sufficiently low luminal pH in conditions like achlorhydria or proton pump inhibitor consumption has been associated with relative bacterial "overgrowth" and increased bacterial diversity associated with chronic disease states including gastric cancer.
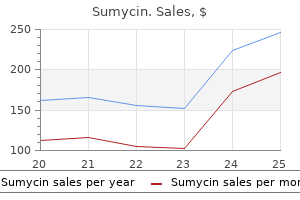
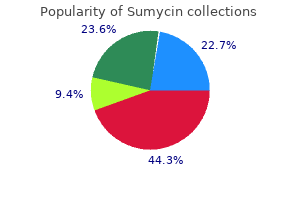
Discount sumycin 250 mg without a prescription. SnapLock® Dance Floors -World's Leading Portable Dance Flooring Company - SnapLock.com.
The lymphoma cells proliferate quickly and spread to the lymph nodes and the small intestine antibiotic resistant uti in pregnancy purchase sumycin toronto. In more severe cases antibiotics for cats order sumycin 500 mg with amex, the lymphoma cells can invade the central nervous system antibiotic resistance nz purchase 250 mg sumycin fast delivery, bone marrow antibiotic for sinus infection chronic sumycin 500 mg buy without prescription, and blood infection under eye 250 mg sumycin buy with visa. If untreated, the disease is fatal, but treatment, especially in the early stages of the disease, has a very good prognosis. The immunosuppressed state of these children leads to recurring bacterial infections, hemorrhage, and death at an early age. Most children who survive the first decade of life are stricken with leukemia or lymphoma. The image resembles a "starry sky" due to the presence of an abundance of tingible-body macrophages. This photomicrograph is of the spleen of a patient with peripheral T-cell lymphoma. T cell B cell Thymus B lymphocytes are believed to remain in the bone marrow to become immunologically competent B cells. Bone marrow Lymph node these immunocompetent T and B cells then seed lymphoid tissues, especially the spleen, lymph nodes, and lymphatic nodules, and are capable of becoming activated (mature) and responding to an antigenic challenge. Mature and immunocompetent cells circulate among the various lymphoid tissues, using blood and lymph vessels. T helper cells play a pivotal role in the development and maintenance of the immune response. They interact with antigen-presenting cells and release cytokines, resulting in the generation of plasma cells for the humoral and T killer (cytotoxic) cells for the cell-mediated response. Capsular arteries Red pulp White pulp Vein Artery Splenic vein Splenic sinusoid Trabeculae Splenic artery the spleen cleanses the s blood, eliminates defunct red blood cells, forms T cells and B cells, and in some animals but not humans, stores red blood cells. The former assemble to form pores in the plasma membrane of the transformed cell, and framentin drives the transformed cell into apoptosis. Observe that the simple columnar epithelium (E) contains not only the nuclei (N) of epithelial cells but also dark, dense nuclei of lymphocytes (arrows), some of which are in the process of migrating from the lamina propria (connective tissue) into the lumen of the duodenum. Note also the presence of a lacteal (La), a blindly ending, lymph-filled lymphatic channel unique to the small intestine. These vessels can be recognized by the absence of red blood cells, although nucleated white blood cells may frequently occupy their lumen. Observe that this particular lymphatic nodule presents no germinal center but is composed of several cell types, as recognized by nuclei of various sizes and densities. Note that the lighter staining germinal center (Gc) is surrounded by the corona (Co) of darker staining cells possessing only a limited amount of cytoplasm around a dense nucleus. Germinal centers form in response to an antigenic challenge and are composed of lymphoblasts and plasmablasts, whose nuclei stain much lighter than those of small lymphocytes. The germinal center is the site of production of small lymphocytes that then migrate to the periphery of the lymphatic nodule to form the corona. Lymph nodes are kidney-shaped structures possessing a convex and a concave (hilar) surface. They are invested by a connective tissue capsule (Ca) that sends trabeculae (T) into the substance of the node, thereby subdividing it into incomplete compartments. The compartmentalization is particularly prominent in the cortex (C), the peripheral aspect of the lymph node. This is the region of B lymphocytes, whereas the paracortex is particularly rich in T lymphocytes. The medullary cords are composed of lymphocytes, macrophages, reticular cells, and plasma cells. Lymph enters the lymph node, and as it percolates through sinuses and sinusoids, foreign substances and nonself antigenic elements are removed from it by phagocytic activity of macrophages. Lymph from the subcapsular sinus enters the cortical sinus and then moves into the medullary sinusoids. It is here that lymphocytes also migrate into the sinusoids, leaving the lymph node via the efferent lymph vessels eventually to enter the general circulation. The hilum of the human lymph node displays the collagenous connective tissue capsule (Ca), from which numerous trabeculae (T) enter into the substance of the lymph node. Note that the basic framework of these medullary cords, as well as of the lymph node, is composed of thin reticular fibers (arrows), which are connected to the collagen fiber bundles of the trabeculae and capsule. The cortex of the lymph node is composed of numerous lymphatic nodules, one of which is presented in this photomicrograph. The thin connective tissue capsule (Ca) sends trabeculae (T) into the substance of the lymph node. Observe that the lymphatic nodule possesses a dark staining corona (Co), composed mainly of small lymphocytes (Ly) whose heterochromatic nuclei are responsible for their staining characteristics. The germinal center (Gc) displays numerous cells with lightly staining nuclei, belonging to dendritic reticular cells, plasmablasts, and lymphoblasts. Both T and B lymphocytes populate the medullary cords, since they are in the process of migrating from the paracortex and cortex, respectively. Some of these lymphocytes will leave the lymph node using the sinusoids and efferent lymphatic vessels at the hilum. The vacuolated appearance of these macrophages is indicative of their active phagocytosis of particulate matter. The pharyngeal tonsil, located in the nasopharynx, is a loose aggregate of lymphatic nodules, often displaying germinal centers (Gc). The epithelial lining (E) is pseudostratified ciliated columnar with occasional patches of stratified squamous nonkeratinized epithelium (asterisk). Note that lymphocytes migrate through the epithelium (arrows) to gain access to the nasopharynx. The crypts frequently contain debris (arrow) that consists of decomposing food particles as well as lymphocytes that migrate from the lymphatic nodules through the epithelium to enter the crypts. The deep surface of the palatine tonsil is covered by a thickened connective tissue capsule (Ca). It is believed that antigen-transporting cells are nonphagocytic and that they trap antigens at the site of antigenic invasion and transport them to lymphatic nodules of lymph nodes, where they mature to become dendritic reticular cells. Transport of immune complexes from the subcapsular sinus to lymph node follicles on the surface of nonphagocytic cells, including cells with dendritic morphology. Immediately deep to the capsule (Ca) lies the subcapsular sinus occupied by three lymphocytes, one of which is labeled (L), as well as the process (P) of an antigen-transporting (antigen-presenting) cell, whose cell body (arrowheads) and nucleus are in the cortex, deep to the sinus. It is invested by a thin connective tissue capsule (Ca) that incompletely subdivides the thymus into lobules (Lo) by connective tissue septa (Se). Each lobule possesses a darker staining peripheral cortex (C) and a lighter staining medulla (M). The connective tissue capsule and septa convey blood vessels into the medulla of the thymus. The thymus begins to involute in the postpubescent individual, and the connective tissue septa become infiltrated with adipocytes. The lobule of the thymus presented in this photomicrograph appears to be completely surrounded by connective tissue septa (Se); three-dimensional reconstruction would reveal this lobule to be continuous with surrounding lobules (Lo). The characteristic light patches of the cortex correspond to the high density of epithelial reticular cells and macrophages (arrows). The cortex of the thymus is bounded externally by collagenous connective tissue septa (Se). Since it lies within the abdominal cavity, it is surrounded by a simple squamous epithelium (E). The germinal center is the site of active production of B lymphocytes during an antigenic challenge. Both the marginal zone and the white pulp are populated with numerous macrophages and antigen-presenting cells (arrowheads), in addition to lymphocytes. The connective tissue framework of the spleen is demonstrated by the use of silver stain, which precipitates around reticular fibers. The regions between sinusoids are occupied by pulp cords, rich in macrophages, reticular cells, and plasma cells. The principal cell of lymphoid tissue is the lymphocyte, of which there are three categories: null cells, B lymphocytes and T lymphocytes. Additionally, macrophages, reticular cells, plasma cells, dendritic cells, and antigen-presenting cells perform important functions in lymphatic tissue. Epithelium Covered by stratified squamous nonkeratinized epithelium that extends into the tonsillar crypts. Capsule the capsule, usually surrounded by adipose tissue, is composed of dense irregular collagenous connective tissue containing some elastic fibers and smooth muscle. Afferent lymphatic vessels enter the convex aspect; efferent lymphatics and blood vessels pierce the hilum. Capsule Dense, irregular collagenous connective tissue capsule separates the tonsil from the underlying pharyngeal wall musculature. Cortex the cortex of a lymph node is characterized by the presence of lymphatic nodules, which have a dark corona, predominantly occupied by B lymphocytes, and lighter staining germinal centers, housing activated B lymphoblasts, macrophages, and dendritic reticular cells. Subcapsular and cortical sinuses possess lymphocytes, reticular cells, and macrophages. Epithelium For the most part, pseudostratified ciliated columnar epithelium (infiltrated by lymphocytes) covers the free surface as well as the folds that resemble crypts. Capsule the thin capsule, situated deep to the tonsil, provides septa for the tonsil. Paracortex the paracortex is the zone between the cortex and medulla, composed of T lymphocytes. Postcapillary venules, with their characteristic cuboidal endothelium, are present. Medulla the medulla displays connective tissue trabeculae, medullary cords (composed of macrophages, plasma cells, and lymphocytes), and medullary sinusoids lined by discontinuous endothelial cells. Lymphocytes, plasma cells, and macrophages are the common cell types in the lumina of sinusoids. The region of the hilum is distinguished by the thickened capsule and lack of lymphatic nodules. Ducts of the seromucous glands, beneath the capsule, pierce the tonsil to open onto the epithelially covered surface. Epithelium Stratified squamous nonkeratinized epithelium covers the tonsil and extends into the shallow crypts. Reticular Fibers With the use of special stains, such as silver stains, an extensive network of reticular fibers may be demonstrated to constitute the framework of lymph nodes. Reticular Fibers With the use of special stains, an extensive network of reticular fibers, which constitute the framework of the spleen, can be demonstrated. Capsule the capsule, composed predominantly of dense irregular collagenous connective tissue, is significantly thickened at the hilum. The capsule also possesses a small amount of elastic fibers and some smooth muscle cells. It is covered by mesothelium (simple squamous epithelium) but is not surrounded by adipose tissue. Trabeculae, bearing blood vessels, extend from the capsule into the substance of the spleen. Capsule the thin capsule is composed of dense irregular collagenous connective tissue (with some elastic fibers). Interlobular trabeculae extending from the capsule incompletely subdivide the thymus into lobules. White Pulp White pulp is composed of periarterial lymphatic sheaths and lymphatic nodules with germinal centers. Both periarterial lymphatic sheaths (predominantly T lymphocytes) and lymphatic nodules (predominantly B lymphocytes) surround the acentrically located central artery. It is composed of lightly staining epithelial reticular cells, macrophages, and densely packed, darkly staining, small T lymphocytes (thymocytes) responsible for the dark appearance of the cortex. Epithelial reticular cells also surround capillaries, the only blood vessels present in the cortex. Marginal Zone A looser accumulation of lymphocytes, macrophages, and plasma cells are located between white and red pulps. The vascular supply of this zone is provided by capillary loops derived from the central artery. It is occupied by plasma cells, lymphocytes, macrophages, and epithelial reticular cells. Pulp cords are composed of delicate reticular fibers, stellateshaped reticular cells, plasma cells, macrophages, and cells of the circulating blood. Sinusoids are lined by elongated discontinuous endothelial cells surrounded by thickened hoop-like basement membrane in association with reticular fibers. These are pulp arterioles, sheathed arterioles, and terminal arterial capillaries. Convincing evidence to determine whether circulation in the red pulp is open or closed is not available, although, in humans, the open circulation is believed to be the most prevalent. The cortex becomes less dense because its population of lymphocytes and epithelial reticular cells is, to some extent, replaced by fat. Reticular Fibers and Sinusoids the thymus possesses neither reticular fibers nor sinusoids. The endocrine system consists of several glands, isolated groups of cells within certain organs, and individual cells scattered among parenchymal cells of the body. This chapter considers only that part of the endocrine system that is composed of glands. The endocrine glands to be discussed here are the · · · · · pituitary, thyroid, parathyroid, suprarenal glands, and pineal body. T derivates are cholesterol derivatives (aldosterone, cortisol, estrogen, progesterone, and testosterone).
These include interleukin 1 antibiotic resistance join the fight purchase sumycin with visa, which stimulates T helper cells and self-activated macrophages antibiotic 100 mg sumycin 500 mg purchase with mastercard, as well as prostaglandin E2 antibiotic induced c diff purchase 500 mg sumycin amex, which attenuates some immune responses antimicrobial underwear for women trusted sumycin 500 mg. Cytokines antibiotic herbs sumycin 250 mg discount, such as interferon-g, released by other lymphoid cells as well as by macrophages, enhance the phagocytic and cytolytic avidity of macrophages. Lymphocytes the lymphocyte, the principal cell of lymphoid tissue, is a key controller responsible for the proper functioning of the immune system. Lymphocytes may be subdivided, according to function, into three categories: null cells, T lymphocytes, and B lymphocytes. It is the T lymphocytes that participate in the graft rejection phenomenon and in the elimination of virally transformed cells. There are three general categories of T cells: naïve T cells, memory T cells, and effector T cells. The activated naïve T cell enters the cell cycle and forms memory T cells and effector T cells. These cells are long-lived, circulating cells that are added to and increase the number of cells of the original clone. It is this increase in the size of the clone that is responsible for the anamnestic response (a more rapid and more intense secondary response) against another encounter with the same antigen. Frequently, T lymphocytes also assist B lymphocytes to amplify and modulate their immune response. The major interactions among T cells, B cells, and antigen-presenting cells are illustrated in Graphics 9-3 to 9-5. B Lymphocytes (B Cells) B lymphocytes (B cells) are formed and become immunocompetent in the bone marrow (bursa of Fabricius in birds). B cells proliferate during a humoral immune response to form plasma cells and B memory cells. In some instances, binding inactivates the antigen, whereas in others the attachment of antibodies to antigens may enhance phagocytosis (opsonization) or activate the complement cascade, resulting in chemotaxis of neutrophils and, frequently, lysis of the invader. B memory cells are similar to T memory cells in that they are long-lived, circulating cells that are added to and increase the number of cells of the original clone. Thus, it is this increase in the size of the clone that is responsible for the anamnestic response against a subsequent encounter with the same antigen. It may be noted that the lymphoid cells are not arranged in any particular pattern but are scattered in a haphazard manner. Frequently, lymphoid nodules, transitory structures that are a denser aggregation of lymphoid tissue composed mainly of lymphocytes, may be observed. Lymphoid nodules may be primary or secondary, where the secondary lymphoid nodules present the characteristic appearance of a lighter germinal center and a darker, peripherally located corona, indicating activation by antigen. The germinal centers are sites of plasma cell production, whereas the corona is produced by mitosis from existing B lymphocytes. Each lymph node has a dense, irregular, collagenous connective tissue capsule and septa, derived from the capsule, subdividing the cortex into incomplete compartments. Attached to the septa and the internal aspect of the capsule is a network of reticular tissue and associated reticular cells that act as a framework for housing the numerous free and migratory cells, mostly lymphocytes, antigen-presenting cells, and macrophages, occupying the organ. The medullary cords are composed mainly of T cells, B cells, and plasma cells that arise in the cortex and paracortex and migrate into the medulla. T cells and B cells enter the sinusoids and leave the lymph node via efferent lymph vessels. Additional cell components of lymph nodes are macrophages, antigen-presenting cells, and some granulocytes. Aside from functioning in the maintenance and production of immunocompetent cells, lymph nodes also filter lymph. Reticular cells and reticular fibers associated with these sinusoids extend into the pulp cords to contribute to the cell population that consists of macrophages, plasma cells, and extravasated blood cells. A region of smaller sinusoids forms the interface between the white and red pulps, and this interface is known as the marginal zone. Capillaries arising from the central arteries deliver their blood to sinusoids of the marginal zone, which is rich in arterial vessels and avidly phagocytic macrophages. Understanding splenic organization depends on knowing the vascular supply of the spleen. Whether these terminal arterial capillaries drain directly into the sinusoids (closed circulation) or terminate as open-ended vessels in the pulp cords (open circulation) has not been determined conclusively; however, in humans, the open circulation is believed to predominate. Participating in the formation of the tonsillar ring are the · palatine, · pharyngeal, and · lingual tonsils. The tonsils produce antibodies against the numerous antigens and microorganisms that abound in their vicinity. There are additional, smaller tonsils, such as the tubal and lingual tonsils, that function in the same manner. Its principal functions are to filter blood, phagocytose senescent red blood cells and invading microorganisms, supply immunocompetent T and B lymphocytes, and manufacture antibodies. Unlike lymph nodes, the spleen is not divided into cortical and medullary regions, nor is it supplied by afferent lymphatic vessels. Blood vessels enter and leave the spleen at its hilum and travel within the parenchyma via trabeculae derived from its connective tissue capsule. The thymus attains its greatest development shortly after birth, but subsequent to puberty, it begins to involute and becomes infiltrated by adipose tissue; however, even in the adult, the thymus retains its ability to form a reduced number of T lymphocytes. The thin connective tissue capsule of the thymus sends septa deep into the organ, incompletely subdividing it into lobules. The major functions of the thymus are the formation, potentiation, and destruction of T lymphocytes. About 90% of double-positive thymocytes are unable to recognize these complexes, and they undergo apoptosis. Single-positive thymocytes that are unable to attack the self are released from the thymus as naïve T lymphocytes. These naïve T cells migrate to the secondary lymphoid organs to set up clones of T cells. Blood vessels gain entrance to the medulla by traveling in the connective tissue septa, which they exit at the corticomedullary junction, where they provide capillary loops to the cortex. Its clinical signs are asymptomatic initially because the swelling of the liver, spleen, and lymph nodes are not accompanied by pain. Other manifestations include the loss of weight, elevated temperature, diminished appetite, and generalized weakness. Histopathologic characteristics include the presence of Reed-Sternberg cells, easily recognizable by their large size, and the presence of two large, pale, oval nuclei in each cell. Most individuals with this syndrome die in early childhood as a result of uncontrollable infections. Lymph Nodes During Infection In a healthy patient with a normal amount of adipose tissue, the lymph nodes are small, soft structures that cannot be palpated easily. However, during an infection, the regional lymph nodes become enlarged and hard to the touch due to the large number of lymphocytes that are being formed within the node. It is relatively rare in the United States but is more common in Central Africa, where it affects young males infected with the Epstein-Barr virus. Nonsteroid-Based Hormones and Amino Acid Derivatives Nonsteroid-based endocrine hormones and amino acid derivatives bind to receptors (some are G protein linked, and some are catalytic) located on the target cell membrane, activate them, and thus initiate a sequence of intracellular reactions. These may act by · altering the state of an ion channel (opening or closing) or · by activating (or inhibiting) an enzyme or group of enzymes associated with the cytoplasmic aspect of the cell membrane. Opening or closing an ion channel will permit the particular ion to traverse or inhibit the particular ion from traversing the cell membrane, thus altering the membrane potential. Some hormones facilitate the opening of calcium channels; · calcium enters the cell, and three or four calcium ions bind to the protein calmodulin, altering its conformation. Thyroid hormones are unusual among the amino acid derivative and nonsteroid-based hormones, in that they directly enter the nucleus, where they bind with receptor molecules. Steroid-Based Hormones Steroid-based endocrine hormones diffuse into the target cell through the plasma membrane and, once inside the cell, bind to a receptor molecule. All of these glands produce hormones that they secrete into the connective tissue spaces. There are three types of hormones, depending on how far they act from their site of secretion: · those that act on the cell, which releases them (autocrine hormones) · those that act in the immediate vicinity of their secretion (paracrine hormones), and · those that enter the vascular system and find their target cells at a distance from their site of origin (endocrine hormones). This chapter details endocrine hormones (see Tables 10-1 and 10-2), whereas other chapters (nervous tissue, respiratory system, and digestive system) discuss autocrine and paracrine hormones. Hormones, based on their chemical nature, are of three types, nonsteroid, steroid based, and amino acid derivatives. The presence of most hormones also elicits a vascularly mediated negative feedback response, in that subsequent to a desired response, the further production and/or release of that particular hormone is inhibited. Although considerable controversy surrounds the classification of these cells vis-à-vis their function, it is probable that at least six of the seven hormones manufactured by the pars anterior are made by separate cells (see Table 10-1). It is believed that two types of acidophils produce somatotropin and prolactin, whereas various populations of basophils produce the remaining five hormones. They are believed to be acidophils and basophils that have released their granules. Control of Anterior Pituitary Hormone Release: · the axons of parvicellular, hypophyseotropic neurons whose soma are located in the paraventricular and arcuate nuclei of the hypothalamus terminate at the primary capillary bed. These axons store releasing hormones (somatotropinreleasing hormone, prolactin-releasing hormone, corticotropin-releasing hormone, thyrotropin-releasing hormone, and gonadotropin-releasing hormone) and inhibitory hormones (prolactin-inhibiting hormone, inhibin, and somatostatin). The hormones are released by these axons into the primary capillary plexus and are conveyed to the secondary capillary plexus by the hypophyseal portal veins. The hormones then activate (or inhibit) chromophils of the adenohypophysis, causing them to release or prevent them from releasing their hormones. Since the pituitary gland develops from two separate embryonic origins, the epithelium of the pharyngeal roof and the floor of the diencephalon, it is frequently discussed as being subdivided into two parts: · the adenohypophysis (pars anterior, pars tuberalis, and pars intermedia) and the · neurohypophysis (pars nervosa and infundibular stalk). The pars nervosa is continuous with the median eminence of the hypothalamus via the thin neural stalk (infundibular stalk). The pituitary gland receives its blood supply from the right and left superior hypophyseal arteries, serving the median eminence, pars tuberalis, and the infundibulum, and from the right and left inferior hypophyseal arteries, which serve the pars nervosa. Hypophyseal Portal System: the two superior hypophyseal arteries give rise to the · primary capillary plexus located in the region of the median eminence. Pars Anterior the pars anterior is composed of numerous parenchymal cells arranged in thick cords, with large capillaries known as sinusoids, richly vascularizing the intervening regions. The parenchymal cells are classified into two main categories: those whose granules readily take up Pars Intermedia the pars intermedia is not well developed. It is composed of pituicytes, cells believed to be neuroglial in nature that may fulfill a supporting function for the numerous unmyelinated axons of the pars nervosa. The release of these neurosecretory hormones (neurosecretion) is mediated by nerve impulses and occurs at the interface between the axon terminals and the fenestrated capillaries. When the axon is ready to release its secretory products, the pituicytes withdraw their processes and permit the secretory product a clear access to the capillaries. An additional secretory cell type, parafollicular cells (clear cells), is present in the thyroid. They manufacture the hormone calcitonin, which is released directly into the connective tissue in the immediate vicinity of capillaries. Calcitonin (see Table 10-2) helps control calcium concentrations in the blood by inhibiting bone resorption by osteoclasts. Pars Tuberalis the pars tuberalis is composed of numerous cuboidal cells whose function is not known. Parathyroid Glands the parathyroid glands, usually four in number, are embedded in the fascial sheath of the posterior aspect of the thyroid gland. They possess slender connective tissue capsules from which septa are derived to penetrate the glands and convey a vascular supply to the interior. In the adult, two types of parenchymal cells are present in the parathyroid glands: · numerous small chief cells and a smaller number of · large acidophilic cells, the oxyphils. It is enveloped by a connective tissue capsule whose septa penetrate the substance of the gland, forming not only its supporting framework but also its conduit for its rich vascular supply. The parenchymal cells of the gland are arranged in numerous follicles, composed of a simple cuboidal epithelium lining a central colloid-filled lumen. The colloid, secreted and resorbed by the follicular cells, is composed of thyroid hormone that is bound to a large protein, and the complex is known as thyroglobulin. To synthesize thyroid hormone · Iodide from the bloodstream is actively transported into follicular cells at their basal aspect via iodide pumps. These glucocorticoids regulate carbohydrate metabolism, facilitate the catabolism of fats and proteins, exhibit anti-inflammatory activity, and suppress the immune response. Zona reticularis cells secrete weak androgens that promote masculine characteristics. Medulla Parenchymal cells of the medulla, derived from neural crest material, are disposed in irregularly arranged short cords surrounded by capillary networks. They contain numerous granules that stain intensely when the freshly cut tissue is exposed to chromium salts. This is referred to as the chromaffin reaction, and the cells are called chromaffin cells. There are two populations of chromaffin cells that secrete the two hormones (see Table 10-2) of the suprarenal medulla, mainly · epinephrine (adrenaline) or · norepinephrine (noradrenaline). Secretion of these two catecholamines is directly regulated by preganglionic fibers of the sympathetic nervous system that impinge on the postganglionic sympathetic neuron-like chromaffin cells, which are considered to be related to postganglionic sympathetic neurons (see Graphic 10-3). Moreover, scattered, large postganglionic sympathetic ganglion cells in the medulla act on smooth muscle cells of the medullary veins, thus controlling blood flow in the cortex. Suprarenal Glands the suprarenal glands (adrenal glands in some animals) are invested by a connective tissue capsule (see Table 10-2 and Graphics 10-2 and 10-3). The glands are derived from two different embryonic origins, namely, mesodermal epithelium, which gives rise to the cortex, and neuroectoderm, from which the medulla originates. The rich vascular supply of the gland is conveyed to the interior in connective tissue elements derived from the capsule.
References
- Gerber DA, Green M, Jaffe R, et al. Cryptosporidial infections after solid organ transplantation in children. Pediatr Transplant. 2000;4:50-55.
- Matthaiou DK, De Waele J, Dimopoulos G. What is new in the use of aminoglycosides in critically ill patients? Intensive Care Med. 2014;40(10):1553-1555.
- Laszik Z, Mitro A, Taylor FB Jr, et al: Human protein C receptor is present primarily on endothelium of large blood vessels: implications for the control of the protein C pathway, Circulation 96:3633-3640, 1997.
- Liao Y, Yang ZL, Peng JS, et al. Neoadjuvant chemotherapy for gastric cancer: a meta-analysis of randomized, controlled trials. J Gastroenterol Hepatol 2013;28(5):777-782.
- Kin H, Zhao ZQ, Sun HY, et al: Postconditioning attenuates myocardial ischemiareperfusion injury by inhibiting events in the early minutes of reperfusion. Cardiovasc Res 2004;62:74-85.
- Mazzone M, Dettori D, Leite de Oliveira R, et al. Heterozygous deficiency of PHD2 restores tumor oxygenation and inhibits metastasis via endothelial normalization. Cell. 2009;136: 839-851.
- Pareek M, Watson JP, Ormerod LP, et al. Screening of immigrants in the UK for imported latent tuberculosis: a multicentre cohort study and cost-effectiveness analysis. Lancet Infect Dis 2011; 11: 435-444.