

Cymbalta
| Contato
Página Inicial
Emily Greenlee, MD
- Clinical Assistant Professor of Ophthalmology
- Department of Ophthalmology
- Roy J. and Lucille A. Carver College of Medicine
- University of Iowa
- Iowa City, Iowa
Epidermal growth factor receptors in cancer tissues of esophagus, lung, pancreas, colorectum, breast and stomach anxiety symptoms stomach pain 20 mg cymbalta purchase amex. Differential response to keratinocyte growth factor receptor and epidermal growth factor receptor ligands of proliferating and differentiating intestinal epithelial cells anxiety symptoms 0f discount cymbalta 40 mg mastercard. Developmental regulation of epidermal growth factor receptor kinase in rat intestine anxiety untreated cheap cymbalta online master card. Receptor-mediated vectorial transcytosis of epidermal growth factor by Madin-Darby canine kidney cells anxiety treatment cheap cymbalta express. Epidermal growth factor binding, stimulation of phosphorylation, and inhibition of gluconeogenesis in rat proximal tubule anxiety symptoms relief buy cheapest cymbalta. The dynamic expression of the epidermal growth factor receptor and epidermal growth factor ligand family in a differentiating intestinal epithelial cell line. The epidermal growth factor receptor juxtamembrane domain has multiple basolateral plasma membrane localization determinants, including a dominant signal with a polyproline core. Regulation of Caco-2 cell proliferation by basolateral membrane epidermal growth factor receptors. Enhanced expression of epidermal growth factor receptor correlates with alterations of chromosome 7 in human pancreatic cancer. Binding and processing of epidermal growth factor in Panc-I human pancreatic carcinoma cells. The correlation between cytoplasmic overexpression of epidermal growth factor receptor and tumor aggressiveness: poor prognosis in patients with pancreatic ductal adenocarcinoma. Expression of transforming growth factor alpha and epidermal growth factor receptor in human hepatocellular carcinoma. Hepatocyte growth factor, transforming growth factor alpha, and their receptors as combined markers of prognosis in hepatocellular carcinoma. Interaction between epidermal growth factor and its receptor in progression of human gastric carcinoma. Immunohistochemical study of epidermal growth factor and epidermal growth factor receptor in gastric carcinoma. Interrelationship between transforming growth factor-alpha and epidermal growth factor receptor in advanced gastric cancer. Expression of epidermal growth factor receptor in normal colonic mucosa and in adenocarcinomas of the colon. Level and function of epidermal growth factor receptor predict the metastatic potential of human colon carcinoma cells. ErbB-2 the preferred heterodimerization partner of all ErbB receptors, is a mediator of lateral signaling. The type 1 growth factor receptor family: new ligands and receptors and their role in breast cancer. Kapitanovic S, Radosevic S, Slade N, Kapitanovic M, Andelinovic S, Ferencic Z, et al. Expression of erbB-3 protein in colorectal adenocarcinoma: correlation with poor survival. Her-2/neu expression and gene amplification in gastrinomas: correlations with tumor biology, growth, and aggressiveness. Gene amplification and protein overexpression of c-erb-b2 in Barrett carcinoma and its precursor lesions. Identification of discriminators of hepatoma by gene expression profiling using a minimal dataset approach. Characterization of a naturally occurring ErbB4 isoform that does not bind or activate phosphatidyl inositol 3-kinase. Ectodomain cleavage of ErbB-4: characterization of the cleavage site and m80 fragment. Gamma-secretase cleavage and nuclear localization of ErbB-4 receptor tyrosine kinase. A natural ErbB4 isoform that does not activate phosphoinositide 3-kinase mediates proliferation but not survival or chemotaxis. Investigation of the prognostic value of coexpressed erbB family members for the survival of colorectal cancer patients after curative surgery. Importance of epidermal growth factor receptor signaling in establishment of adenomas and maintenance of carcinomas during intestinal tumorigenesis. Mice harboring a defective epidermal growth factor receptor (waved-2) have an increased susceptibility to acute dextran sulfateinduced colitis. Aberrant neural and cardiac development in mice lacking the ErbB4 neuregulin receptor. Colonic epithelial expression of ErbB2 is required for postnatal maintenance of the enteric nervous system. Employment of the epidermal growth factor receptor in growth factor-independent signaling pathways. Phosphorylation of beta-catenin and epidermal growth factor receptor by intestinal trefoil factor. Expression of somatomedin/insulin-like growth factor messenger ribonucleic acids in the human fetus: identification, characterization, and tissue distribution. Insulin/insulin-like growth factor I hybrid receptors have different biological characteristics depending on the insulin receptor isoform involved. Insulin-like growth factor I rapidly stimulates tyrosine phosphorylation of a Mr 185,000 protein in intact cells. Coupling of the insulin-like growth factor-I receptor tyrosine kinase to Gi2 in human intestinal smooth muscle: G-dependent mitogen-activated protein kinase acitvation and growth. Demonstration of tumor suppression by mannose 6-phosphate/insulin-like growth factor 2 receptor. Insulin-like growth factor-binding protein-5 stimulates growth of human intestinal muscle cells by activation of Gi3. Nuclear import of insulin-like growth factor-binding protein-3 and -5 is mediated by the importin beta subunit. Insulinlole growth factor binding protein-5 interacts with the vitamin D receptor and modulates the vitamin D response in osteoblasts. Determination of the histological distribution of insulin like growth factor 1 receptors in the rat gut. Tissue and development specific regulation of a complex family of rat insulin-like growth factor I messenger ribonucleic acids. Insulin-like growth factor I messenger ribonucleic acids with alternative 5-untranslated regions are differentially expressed during development of the rat. Relative expression and localization of the insulin-like growth factor system components in the fetal, child and adult intestine. Expression of two types of receptor for insulinlike growth factors in human colonic epithelium. Regulation and localization of the insulin-like growth factor system in small bowel during altered nutrient status. Expression and regulation of the insulin-like growth factor axis components in rat liver myofibroblasts. Insulin-like growth factors and the developing and mature rat small intestine: receptors and biological actions. Insulin-like growth factor-I and the gastrointestinal system: therapeutic indications and safety implications. Modulation of epidermal growth factor-induced cell proliferation and receptor binding by insulin in cultured intestinal epithelial cells. Epidermal growth factor primes intestinal epithelial cells for proliferative effect of insulinlike growth factor I. Extracellular thiol/ disulfide redox state affects proliferation rate in a human colon carcinoma (Caco2) cell line. Insulin-like growth factor signaling pathways in rat hepatic stellate cells: importance for deoxyribonucleic acid synthesis and hepatocyte growth factor production. Regulation of gastrointestinal growth in fetal sheep by luminally administered insulin-like growth factor-I. Stimulation and inhibition of proliferation in the small intestinal crypts of the mouse after in vivo administration of growth factors. Truncated and native insulinlike growth factor I enhance mucosal adaptation after jejunoileal resection. Beneficial effects of insulin-like growth factor I on epithelial structure and function in parenterally fed rat jejunum. Insulin-like growth factor 1 has beneficial effects, whereas growth hormone has limited effects on postoperative protein metabolism, gut integrity, and splenic weight in rats with chronic mild liver injury. Insulin-like growth factor-I prevents gut atrophy and maintains intestinal integrity in septic rats. Enhanced growth of small bowel in transgenic mice overexpressing bovine growth hormone. Enhanced growth of small bowel in transgenic mice expressing human insulin-like growth factor I. Lowdose growth hormone in adult home parenteral nutrition-dependent short bowel syndrome patients: a positive study. Suppressor of cytokine signaling-2: a growth hormone-inducible inhibitor of intestinal epithelial cell proliferation. Insulin-like growth factor 1 inhibits apoptosis using the phosphatidylinositol 3-kinase and mitogen-activated protein kinase pathways. Growth factors in inflammatory bowel disease: the actions and interactions of growth hormone and insulin-like growth factor-I. Expression of insulin-like growth factor I by activated hepatic stellate cells reduces fibrogenesis and enhances regeneration after liver injury. Minireview: tissue-specific versus generalized gene targeting of the igf1 and igf1r genes and their roles in insulin-like growth factor physiology. Mechanisms of regulatory peptide action in the gastrointestinal tract: trefoil peptides. Pharmacology of a new porcine pancreatic polypeptide with spasmolytic and gastric acid secretion inhibitory effects. Dermal glands of Xenopus laevis contain a polypeptide with a highly repetitive amino acid sequence. Distinct pathways of cell migration and antiapoptotic response to epithelial injury: structure-function analysis of human intestinal trefoil factor. Transgenic mice that overexpress the human trefoil peptide pS2 have an increased resistance to intestinal damage. Crystal structure of a disulfide-linked "trefoil" motif found in a large family of putative growth factors. Identification and characterization of rat intestinal trefoil factor: tissue- and cell-specific member of the trefoil protein family. Cloning of contiguous genomic fragments from human chromosome 21 harbouring three trefoil peptide genes. The gene encoding mouse intestinal trefoil factor: structural organization, partial sequence analysis, and mapping to murne chromosome 17q. Gastroprotective peptide trefoil factor family 2 gene is activated by upstream stimulating factor but not by c-Myc in gastrointestinal cancer cells. Estrogen-responsive element of the human pS2 gene is an imperfectly palindromic sequence. Characterization of the genomic structure and the promoter region of the human intestinal trefoil factor. Hypoxia-inducible factor 1-dependent induction of intestinal trefoil factor protects barrier function during hypoxia. Identification of a goblet cellspecific enhancer element in the rat intestinal trefoil factor gene promoter bound by a goblet cell nuclear protein. Goblet-cell-specific transcription of mouse intestinal trefoil factor gene results from collaboration of complex series of positive and negative regulatory elements. Keratinocyte growth factor promotes goblet cell differentiation through regulation of goblet cell silencer inhibitor. A silencer inhibitor confers specific expression of intestinal trefoil factor in gobletlike cell lines. Trefoil peptide expression and secretion is regulated by neuropeptides and acetylcholine. Short-chain fatty acids inhibit intestinal trefoil factor gene expression in colon cancer cells. Mouse trefoil factor genes: genomic organization, sequences and methylation analyses. Helicobacter pylori infection methylates and silences trefoil factor 2 leading to gastric tumor development in mice and humans. The P-domain or trefoil motif: a role in renewal and pathology of mucous epithelia. Trefoil peptides: a newly recognized family of epithelial mucin-associated molecules. Human intestinal trefoil factor is expressed in human hypothalamus and pituitary: evidence for a novel neuropeptide. Estimation of pS2 protein level in human body fluids by a sensitive two-site enzyme immunoassay. Identification and characterization of a novel gastric peptide hormone: the motilin-related peptide. Breast cancer-associated pS2 protein: synthesis and secretion by normal stomach mucosa. Spasmolytic polypeptide is a major antral peptide: distribution of the trefoil peptides human spasmolytic polypeptide and pS2 in the stomach.
Sutures and fontanelles allow the bones of the skull to overlap (molding) during birth anxiety high blood pressure best cymbalta 40 mg. Soon after birth, membranous bones move back to their original positions, and the skull appears large and round anxiety videos order cymbalta 20 mg with amex. Several sutures and fontanelles remain m em branous for a considerable time after birth, which allows bones of the vault to continué to grow after birth to accommodate postnatal growth of the brain anxiety fear cheap cymbalta 30 mg buy on-line. Although a 5- to 7-year-old child has nearly all of his or her cranial capacity, some su tures remain open until adulthood anxiety girl cheap cymbalta master card. Mesenchyme for these structures is derivad from neural crest [blue], paraxial mesoderm [somites and somitomeres] [red], and lateral píate mesoderm lyellow] anxiety symptoms 5 yr old 40 mg cymbalta buy visa. Neurocranium the neurocranium is m ost conveniently divided into two portions: (1) the membranous part, consisting of flat bones, which surround the brain as a vault, and (2) the cartilaginous part, or chondrocranium, which forms bones of the base of the skull. Mesenchyme from these two sources invests the brain and undergoes intram em branous ossification. The posterior fontanelle closes about 3 m onths after birth; the anterior fontanelle Gloses around the middie of the second year. Those that lie in front of the rostral lim it of the notochord, which ends at the level of the pituitary gland in the center of the sella turcica, are derived from neural crest cells. Those that he posterior to this lim it arise from occipital sclerotomes formed by paraxial m esoderm and form the chordal chondrocranium. The base of the skull is form ed when these cartilages fuse and ossify by endochondral ossification. Those form ing posterior to this landmark arise from paraxial mesoderm [chordal chondrocranium) [red]. Víscerocranium the viscerocranium, which consists of the bones of the face, is form ed m ainly from the first two pharyngeal arches (see Chapter 17). The first arch gives rise to a dorsal portion, the m axillary process, which extends forward beneath the región of the eye and gives rise to the m axilla, the zygom atic bone, and p art of the tem poral bone. M esenchym e around the M eckel cartilage condenses and ossifies by intram em branous ossification to give rise to the m andible. The M eckel cartilage whether ossification of the skull is proceeding normally and whether intracranial pressure is normal. In most cases, the anterior fontanelle closes by 18 months of age, and the posterior fontanelle closes by 1 to 2 months of age. The dorsal tip of the m andibular process, along with that of the second pharyngeal arch, later gives rise to the in cu s, the m alleus, and the stapes. O ssification of the three ossicles begins in the fourth m onth, m aking these the first bones to becom e fully ossified. M esenchym e for form ation of the bones of the face is derived from neural crest cells, including the nasal and lacrim al bones. This appearance is caused by (1) virtual absence of the paranasal air sinuses and (2) the small size of the bones, particularly the jaws. With the appearance of teeth and development of the air sinuses, the face loses its babyish characteristics. Neural Crest Cells Neural crest celis originating in th e neuroectoderm fo rm th e facial skeleton and part of the skull. Cranial neural folds fail to elevate and fuse, leaving the cranial neuropore open. Note the long narrow shape of the head with prom inent frontal and occipital regions. Child with plagiocephaly resulting from premature closure of the coronal suture on one side of the skull. O ther parts of the skeleton are affected as w ell and often the clavicles are underdeveloped or missing, as in this case. In m ost cases, microcephaly is associated with significant intellectual disabilities. A typical vertebra consists of a vertebral arch and foram en (through which the spinal cord passes), a body, transverse processes, and usually a spinous process. Sclerotome cells are dispersing to migrate around the neural tube and notochord to contribute to vertebral form ation. A t the fourth week of development, sclerotom ic segments are separated by less dense intersegmental tissue. Note the position of the myotomes, intersegm ental arteries, and segmental nerves. Proüferation of the caudal half of one sclerotom e proceeds into the intersegm ental mesenchyme and cranial half of the subjacent sclerotome [orrows]. Vertebrae are form ed by the upper and lower halves of tw o successive sclerotom es and the intersegm ental tissue. Myotomes bridge the intervertebral discs, and therefore, can move the vertebral column. As develop ment continúes, the sclerotome portion of each somite also undergoes a process called resegmentatíon. Resegmentation occurs when the caudal half of each sclerotome grows into and fiises with the cephalic half of each subjacent sclerotome arrow s in. As a result of this process, muscles derived from the myotome región of each somite become attached to two adjacent somites across the in tervertebral discs and can therefore move the vertebral column. Mesenchymal cells between cephalic and caudal parts of the original sclerotome segment do not proliferate but fill the space between two precartilaginous vertebral bodies. Although the notochord regresses entirely in the región of the vertebral bodies, it persists and enlarges in the región of the intervertebral disc. Here it contributes to the nucleus pulposus, which is later surrounded by circular fibers of the annulus fíbrosus. Resegmentation of sclerotomes into definitive vertebrae causes the myotomes to bridge the in tervertebral discs, and this alteration gives them the capacity to move the spine. For the same reason, intersegmental arteries, at first lying between the sclerotomes, now pass midway over the vertebral bodies. Spinal nerves, however, come to lie near the intervertebral discs and leave the vertebral column through the intervertebral foramina. As the vertebrae form, two prim ary curves of the spine are established: the thoracic and sacral curvatures. Later, two secondary curves are estab lished: the cervical curvature, as the child learns to hold up his or her head, and the lumbar curvature, which forms when the child learns to walk. Ultrasound sean of a 26-w eekfetus with spina bifida in the lumbosacral región [asterísk]. Because of the shape of the skuli, the image is called the " lemon sign," which occurs in some of these cases and is due to the brain being pulled caudally, changing the shape of the head [see Arnold-Chiari malformation, p. Costal cartilages are formed by sclerotome cells that migrate across the lateral som itic frontier into the adjacent lateral píate mesoderm (see Chapter 11 for a description of the lateral som itic frontier). The sternum develops independently in the parietal layer of lateral píate mesoderm in the ventral body waU. Two sternal bands are formed in the parietal (somatic) layer of lateral píate mesoderm on either side of the midline, and these later fuse to form cartilaginous models of the manubrium, sternebrae, and xiphoid process. Pectus cari- natum refers to a flatten ing of the chest bilaterally with an anteriorly projecting sternum. Both defects m ay result from abnorm alities of ventral body w all closure or form ation of th e costal cartilages and sternum. Some bones, such as the flat bones of the skull, undergo intram em branous ossification; that is, mesenchyme cells are directly transformed into osteoblasts. In most bones, such as the long bones of the limbs, mesenchyme condenses and forms hyaline cartilage models of bones. Ossification centers appear in these cartilage models, and the bone gradually ossifies by endochondral ossification. Neural crest cells form the face, part of the cranial vault, and the prechordal part of the chondrocranium (the part that lies ros tral to the pituitary gland). The vertebral colum n and ribs develop from the sclerotom e com partm ents of the som ites, and the sternum is derived from m esoderm in the ventral body wall. A definitive vertebra is formed by condensation of the caudal h alf of one sclerotom e and fusión with the cranial h alf of the subjacent sclerotom. The many abnormalities of the skeletal sys tem include vertebral (spina bifida), cranial (cranioschisis and craniosynostosis), and facial (cleft palate) defects. M ajor malformations of the limbs are rare, but defects of the radius and digits are often associated with other abnormali ties (syndromes). Skeletal muscle is derived from paraxial mesoderm, which forms somites from the occipital to the sacral regions and somitomeres in the head. Smooth muscle difFerentiates from visceral splanchnic mesoderm surrounding the gut and its derivatives and from ectoderm (pupillary, mammary gland, and sweat gland muscles). Cardiac muscle is derived from visceral splanchnic mesoderm surrounding the heart tube. Here they form infrahyoid, abdominal wall (rectus abdominis, internal and external oblique, and transversus abdominis), and limb muscles. The remaining cells in the myotome form muscles of the back, shoulder girdle, and intercostal muscles (Table 11. Initially, there is a well-defined border between each somite and the parietal layer of lat eral píate mesoderm called the lateral somitic frontier. The prim axial dom ain that comprises the región around the neural tube and contains only somite-derived (paraxial mesoderm) cells 2. Regardless of their domain, each m yotom e receives its innervation from spinal nerves derived from the same segment as the muscle cells. The lateral somitic frontier also defines the border between dermis derived from dermatomes in the back and dermis derived from lateral píate mesoderm in the body wall. Musculature of the axial skeleton, body wall, and limbs is derived from somites, which initially form as somitomeres and extend from the occipital región to the tail bud. Immediately after segmentation, these somitomeres undergo a process of epithelization and form a "ball" of epithelial cells with a small cavity in the center. The ventral región of each somite then becomes mesenchymal again and forms the sclerotome. Cells from these two areas migrate and proliferate to form progenitor muscle cells ventral to the derm atom e, thereby forming the derm om yotom. Cells in the ventral and medial walls of the somite lose their epithelial characteristics and migrate around the neural tube and notochord, and some move into the parietal layer of lateral píate mesoderm. Cells from both regions migrate ventral to the dermatome to form the dermomyotome. In combination, somitic cells and lateral píate mesoderm cells constitute the abaxial mesodermal domain, whereas the primaxial mesodermal domain oniy contains somitic cells [paraxial mesoderm). Together, dermatome cells and the muscle cells that associate with them form the dermomyo tome. The dermomyotome begins to differentiate: Myotome cells contribute to primaxial muscles, and derma tome cells form the dermis of the back. Hypaxial muscles [limb and body wall] are Innervated by ventral (anterior) prim ary rami. The new description is based on the actual embryological origín of muscle cells from two different populations of muscle cell precursors, the abaxial and primaxial cells, and not their innervation. The description does not preclude the fact that epaxial (above the axis) muscles (back mus cles) are innervated by dorsal prim ary rami, whereas hypaxial (below the axis) muscles (body wall and limb muscles) are innervated by ventral prim ary ram i. In the head región, these connective tissues are derived from neural crest cells; in cervical and occipital regions, they differentiate from somitic m esoderm; and in the body wall and limbs, they origínate from the parietal layer of lateral píate mesoderm. Myofibrils soon appear in the cytoplasm, and by the end of the third month, cross-striations, typical of skeletal muscle, ap pear. A similar process occurs in the seven somitomeres in the head región rostral to the occipital somites. However, somitomeres never segregate into recognizable regions of sclerotome and dermomyotome segments prior to difFerentiation. Tendons for the attachment of muscles to bones are derived from sclerotome cells lying adjacent to myotomes at the anterior and posterior borders of somites. Patterns of muscle formation in the head are directed by connective tissue elements derived from neural crest cells. The mesenchyme is derived from dorsolateral cells of the som ites that migrate into the lim b bud to form the muscles. As in other regions, connective tissue dictates the pattern of m uscle form ation, and this tissue is derived from the pa rietal layer of lateral píate m esoderm, which also gives rise to the bones of the lim b (see Chapter 12). In the coronary arteries, smooth muscle originates from proepicardial cells (see Chapter 13) and neural crest cells (proximal segments). Smooth muscle in the wall of the gut and gut derivatives is derived from the splanchnic layer of lateral píate mesoderm that surrounds these structures. Only the sphincter and dilator muscles of the pupil and muscle tissue in the mammary and sweat glands are derived from ectoderm. This factor is upregulated by growth factors through kinase phosphorylation pathways. Myoblasts adhere to one another by special attachments that later develop into intercalated discs. During later development, a few special bundles of muscle cells with irregularly distributed myofibrils become visible. Skeletal muscles are derived from paraxial mesoderm, in cluding (1) somites, which give rise to muscles of the axial skeleton, body wall, and limbs, and (2) somitomeres, which give rise to muscles of the head. This frontier or border separates two mesodermal domains in the embryo: (1) the primaxial domaín that surrounds the neural tube and contains only somite-derived cells (paraxial mesoderm) and (2)the abaxial domain that consists of the parietal layer of lateral píate meso derm in combination with somite-derived cells that migrate across the frontier into this región. Abaxial muscle precursor cells diíferentiate into infrahyoid, abdominal wall (rectus abdominus, extem al and internal obliques, transversus abdominus), and limb muscles. Primaxial muscle precursor cells form muscles of the back, some muscles of the shoulder girdle, and intercostal muscles (Table 11.
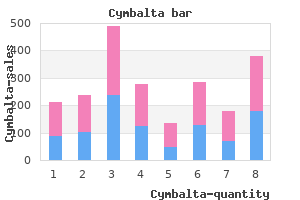
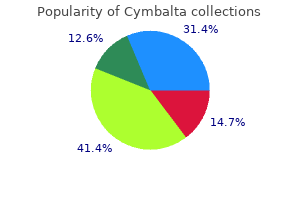
40 mg cymbalta purchase amex. LIVING WITH ANXIETY/PANIC DISORDER My genuine experience at Beat The Bomb!!!!.
For example, the colon of neonates has villous-like structures426,427 and active transport of some dietary components that are lacking in the colon of adults anxiety symptoms only at night 40 mg cymbalta buy with amex. Furthermore, transport functions also vary along the length of the small intestine during postnatal development anxiety 7dpo discount cymbalta 40 mg without a prescription. Other notable differences are the distribution and presence of vacuolated enterocytes and saturable absorption of bile acids and some vitamins, all of which are present only in early neonatal development anxiety side effects purchase cymbalta on line amex. There are also distinct changes in transport function along the crypt villus axis noted during development anxiety breathing techniques buy 60 mg cymbalta with amex. This latter possibility seems particularly relevant as cell replacement and mitotic activity are slower in the perinatal period, and increase around the suckling/ weaning transition anxiety symptoms in 9 year old boy order discount cymbalta on-line. Several physiologic factors that influence intestinal transport function are shown. Luminal effectors, paracrine effectors, nervous stimuli, and hormonal secretions all play a role in regulating transport function during postnatal development. M cells have a modified basolateral surface that forms an invagination into which immune cells can migrate. Enteroendocrine cells, lymphocytes, and other immune cells respond to stimuli by the release of signaling mediators that influence other cell types of the lamina propria through paracrine action. The enteric nervous system receives its input from paracrine effectors, from sensory neurons, and from the central nervous system. This information is processed in the enteric nervous plexus, which results in the activation of motor neurons that may control various mucosal functions. Lastly, hormonal or endocrine stimuli influence the cells of lamina propria, as well as cells of the epithelial layer. Administration of glucocorticoid to suckling animals has broad effects on many digestive functions, including sucrase, alkaline phosphatase, enteropeptidase, diamine oxidase, pepsin, and amylase expression. These hormones bind to specific intracellular receptors (glucocorticoid receptors). The hormone/receptor complex undergoes conformational change and activates a transcriptional complex to bind to hormone-specific ciselements on gene promoters or enhancers, thus inducing expression of genes that control intestinal development. Food processed in the gastrointestinal tract induces a series of physiologic responses, including the release of trophic hormones; the stimulation of the enteric nervous system; and the activation of secretary, digestive, and absorptive functions. Because the small intestine is directly exposed to environmental factors as the result of daily dietary intake, it is prepared to adapt its structure and function in response to variations in the diet. The normal diet also contains growth factors that directly stimulate the growth of intestinal mucosal cells. The developmental patterns of the gastrointestinal tract at these stages are controlled by both genetic and dietary factors. In some species, the neonatal intestine undergoes rapid growth and reorganization, possibly as a result of programmed gene expression associated with the immediate response to milk. The milk of mammals contains a range of trophic factors, hormones, and other biologically active substances that may cause growth-enhancing effects on the gastrointestinal mucosa. As a consequence of the rumen maturation, which coincides with weaning, dietary carbohydrates are fermented into short chain fatty acids by rumen microflora, the amount of hexoses entering the small intestine gradually decreases,477 and the capacity of Na-glucose cotransport declines. However, the ontogenic decline in lactase activity can be promoted or delayed by changing the time when rats are switched from milk to the adulttype diet. Although the ontogenic decline in intestinal lactase is intrinsically programmed, luminal factors modulate this process. The transition to the adult diet is required for the normal progression of events. Changes in small intestinal mucosa morphology and cell renewal in suckling, prolonged-suckling, and weaned lambs. Cell migration pathway in the intestinal epithelium: an in situ marker system using mouse aggregation chimeras. Intestinal epithelial cell differentiation: new insights from mice, flies and nematodes. Chapter 14 Molecular Physiology of Gastrointestinal Function during Development 437 12. Continuous gastric pH measurement in young and older healthy preterm infants receiving formula and clear liquid feedings. Parietal cell function of full-term and premature infants: unstimulated gastric acid and intrinsic factor secretion. High prevalence of asymptomatic esophageal and gastric lesions in preterm infants in intensive care. Gastric H(),K()-adenosine triphosphatase beta subunit is required for normal function, development, and membrane structure of mouse parietal cells. Alterations in gastric mucosal lineages induced by acute oxyntic atrophy in wild-type and gastrin-deficient mice. Gastric and duodenal mucosal bicarbonate secretion, in Physiology of the gastrointestinal tract. Effect of age on the secretory capacity of pig small intestine in vivo and in vitro. L-glutamine with D-glucose stimulates oxidative metabolism and NaCl absorption in piglet jejunum. Taurodeoxycholate and the developing rabbit distal colon: absence of secretory effect. Immunohistochemical localization of cystic fibrosis transmembrane conductance regulator in human fetal airway and digestive mucosa. Chloride transport in primary cultures of rabbit colonocytes at different stages of development. Electrophysiological characterization of chloride secretion across the jejunum and colon of pigs as affected by age and weaning. Age and cortisone alter host responsiveness to cholera toxin in the developing gut. The immature rat small intestine exhibits an increased sensitivity and response to Escherichia coli heat-stable enterotoxin. Age-related differences in receptors for Escherichia coli heat-stable enterotoxin in the small and large intestine of children. Mechanisms of increased susceptibility of immature and weaned pigs to Escherichia coli heat-stable enterotoxin. Novel sites for expression of an Escherichia coli heat-stable enterotoxin receptor in the developing rat. Development of intestinal host defense: an increased sensitivity in the adenylate cyclase response to cholera toxin in suckling rats. Cholera enterotoxin-induced mucus secretion and increase in the mucus blanket of the rabbit ileum in vivo. Involvement of 5-hydroxytryptamine, prostaglandin E2, and cyclic adenosine monophosphate in cholera toxininduced fluid secretion in the small intestine of the rat in vivo. Actions of serotonin antagonists on cholera-toxin-induced intestinal fluid secretion. Neural mediation of cholera toxin-induced mucin secretion in the rat small intestine. Developmental pattern of small intestinal enterokinase and disaccharidase activities in the human fetus. Intestinal disaccharidase activities in relation to age, race, and mucosal damage. An upstream polymorphism associated with lactase persistence has increased enhancer activity. Differential distribution of digestive enzymes in isolated epithelial cells from developing human fetal small intestine and colon. Transient coappearance of glucagon and insulin in the progenitor cells of the rat pancreatic islets. Immunocytochemical studies on pancreatic endocrine cells at early stages of development of the pig. Precursor cells of mouse endocrine pancreas coexpress insulin, glucagon and the neuronal proteins tyrosine hydroxylase and neuropeptide Y, but not pancreatic polypeptide. The Pax4 gene is essential for differentiation of insulin-producing beta cells in the mammalian pancreas. Pax6 is required for differentiation of glucagonproducing alpha-cells in mouse pancreas. Essential requirement for Pax6 in control of enteroendocrine proglucagon gene transcription. Expression of adrenomedullin and its receptor during embryogenesis suggests autocrine or paracrine modes of action. Expression pattern for adrenomedullin during pancreatic development in the rat reveals a common precursor with other endocrine cell types. A newly discovered role of transcription factors involved in pancreas development and the pathogenesis of diabetes mellitus. Pancreatic transcription factors and their role in the birth, life and survival of the pancreatic beta cell. Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Changes in fatty acid composition during cell differentiation in the small intestine of suckling piglets. Changes in phospholipid and cholesterol concentrations of the rat microvillus membrane during maturation. Ontogeny of basolateral membrane lipid composition and fluidity in small intestine. Lipid composition and membrane fluidity in the small intestine of the developing rabbit. Chapter 14 Molecular Physiology of Gastrointestinal Function during Development 439 94. Dietary triacylglycerol modulates sodiumdependent D-glucose transport, fluidity and fatty acid composition of rat small intestinal brush-border membrane. The lipid fluidity of rat colonic brush-border membrane vesicles modulates Na-H exchange and osmotic water permeability. Morphometrical changes in the apical surface of the colonic absorptive cells in perinatal rats with special reference to the effect of fetal oral administration of milk in utero. The development of gut associated lymphoid tissue in the terminal ileum of fetal human intestine. Cessation of uptake of macromolecules by neonatal guinea pig, hamster and rabbit intestinal epithelium (closure) and transport into blood. Development of the neonatal rat small intestinal barrier to nonspecific macromolecular absorption. Decrease in intestinal permeability to polyethylene glycol 1000 during development in the pig. Hormonal control of intestinal Fc receptor gene expression and immunoglobulin transport in suckling rats. Intestinal macromolecular transmission in the young rat: influence of protease inhibitors during development. Membrane-bound and fluid-phase macromolecules enter separate prelysosomal compartments in absorptive cells of suckling rat ileum. Proteolytic activity as a regulator of the transmission of orally fed proteins from the gut to the blood serum in the suckling rat. Development of dome epithelium in gutassociated lymphoid tissues: association of IgA with M cells. Role of passive and adaptive immunity in influencing enterocyte-specific gene expression. IgG binding and expression of its receptor in rat intestine during postnatal development. Transepithelial transport of maternal antibody: purification of IgG receptor from newborn rat intestine. Binding of subclasses of rat immunoglobulin G to detergent-isolated Fc receptor from neonatal rat intestine. Distinction between jejunal and ileal epithelial cells demonstrated by simultaneous ultrastructural localization of IgG and acid phosphatase. The effect of inhibition of proteolysis on the uptake of macromolecules by the intestine of the newborn rabbit before and after weaning. Characterization of the rat intestinal Fc receptor (FcRn) promoter: transcriptional regulation of FcRn gene by the Sp family of transcription factors. Sp1 and kruppel-like factor family of transcription factors in cell growth regulation and cancer. Transcriptional control of the murine polymeric IgA receptor promoter by glucocorticoids. Stability and distribution of orally administered epidermal growth factor in neonatal pigs. Expression of insulin receptors and of 60-kDa receptor substrate in rat mature and immature enterocytes. Characterization and autoradiographic localization of the epidermal growth factor receptor in the jejunum of neonatal and weaned pigs. Specific receptors for epidermal growth factor in rat intestinal microvillus membranes. Nutrient-independent and nutrient-dependent factors stimulate protein synthesis in colostrum-fed newborn pigs. Small intestinal disaccharidase activity and ileal villus height are increased in piglets consuming formula containing recombinant human insulin-like growth factor-I. Growth of artificially fed infant rats: effect of supplementation with insulin-like growth factor I. Uptake and transepithelial transport of nerve growth factor in suckling rat ileum. Ontogenetic development of monosaccharide and amino acid transporters in rabbit intestine.
The suprafacial zone has the following boundaries: laterally, the superior cerebellar peduncle; medially, a vertical line 2 mm lateral to the median sulcus; superiorly, the frenulum veli; and inferiorly, the facial colliculus anxiety symptoms rash cymbalta 20 mg purchase without prescription. In such cases, the facial colliculus can be stimulated directly while the facial nerve is monitored anxiety symptoms dry mouth 30 mg cymbalta order visa. Typically, however, lesions of the floor of the fourth ventricle are not treated surgically unless the patient has a complete neurological deficit such as internuclear ophthalmoplegia, abducens palsy, or facial paresis anxiety 6 things you can touch with your hands discount cymbalta 40 mg without a prescription. At the end of the procedure, the dura and fascia are closed in layers and the bone flap is replaced anxiety tremors order cymbalta visa. Otherwise, significant spasms can occur and preclude adequate reapproximation of the suboccipital musculature to the occiput anxiety symptoms fever discount cymbalta 30 mg online. The technique, as performed at our institution, has been detailed elsewhere25 and is reviewed here briefly. Slight extension of the neck, so that the malar eminence is the most superior point in the operative field, causes the frontal lobe to fall away from the anterior cranial fossa. The scalp incision extends from the root of the zygoma, 1 cm anterior to the tragus, to the midline or medial contralateral line. A vascularized pericranial flap should be preserved during the opening for use at the end of the procedure to isolate the exenterated frontal sinus should it be violated during the osteotomy. The temporalis fascia is incised along the posterior border of the skin incision and extended anteriorly just below the superior temporal line. A dissection plane is then developed between the temporalis fascia and muscle, below the second fat pad. Next, the frontozygomatic and temporal zygomatic processes and superior orbital rim are exposed by elevating the temporalis fascia off the respective bony surfaces with a periosteal elevator. The elevator should not be passed too deeply toward the cone of the orbit because the optic nerve can be damaged. After the temporalis muscle is elevated and retracted from the squamous temporal bone, a standard pterional craniotomy is performed, followed by the orbitozygomatic osteotomy. The temporalis muscle is again released and placed over the craniectomy defect in preparation for the orbitozygomatic osteotomy. For the conventional orbitozygomatic approach, an oscillating saw is used to make six osteotomies. The first osteotomy is made at the base of the zygoma by placing the saw on the zygomatic root just above the temporalis muscle. It must not be placed too deep to avoid violating the capsule of the temporomandibular joint. The second cut begins just anterior and inferior to the temporal process of the zygomatic bone and proceeds laterally to medially while directed slightly inferiorly. It stops in the midportion of the malar eminence at the zygomaticofacial foramen, which is connected with the third cut. This latter cut extends from the inferior orbital fissure from within the orbit through the orbital surface of the temporal bone and connects the second cut at an apex. An inverted V is thus created on the malar eminence; the left and right links are the second and third cuts, respectively. The fourth cut extends through the orbital surface of the frontal bone posteriorly toward the superior orbital fissure. If more medial access is required, the supraorbital nerve can be mobilized from its foramen with an osteotome. If the foramen is high above the orbital rim, the nerve can be sacrificed, but forehead numbness will result. It extends posteriorly from the inferior orbital fissure across the greater wing of the sphenoid bone and through the posterior orbit. Alternatively, it can connect the fourth and fifth cuts just proximal to the superior orbital fissure. A rongeur can be used to remove the residual bony island of the greater wing of the sphenoid until the dural fold of the superior orbital fissure is identified. Importantly, the orbitozygomatic approach can be tailored to suit individual circumstances (modified orbitozygomatic approach) when a lesion requires less rostral visualization. However, for most deep-seated lesions, complete removal of the orbital roof and zygoma is indicated. To ensure precise reapproximation at closure, holes can be drilled for a cranial fixation plate before the osteotomy is removed. Tacking sutures are placed deep toward the orbital apex and anchored around the secure fishhooks. The operating microscope is then brought into the field, and a transsylvian or subtemporal approach is undertaken in atraumatic fashion. The working distance to lesions in the parasellar region and the interpeduncular fossa is about 3 cm shorter with an orbitozygomatic approach than with a standard frontotemporal approach. A more upward and oblique view of the sylvian fissure, third ventricle, and upper brainstem can be achieved with less retraction on the temporal and frontal lobes. Patients should be forewarned of significant postoperative periorbital edema and diplopia, which usually resolve within a week. On postoperative day 1, patients can begin jaw exercises to avoid restricted range of motion of the temporomandibular joint as a result of scarring of the temporomandibular ligament and joint capsule. Atrophy of the temporalis muscle can be minimized by using monopolar cauterization judiciously and by reapproximating the superior aspect of the temporalis muscle to a small muscle cuff of fascia left along the superior temporal line. The patient can be positioned lateral, prone, or supine with a sandbag beneath the ipsilateral shoulder. In the supine position, the head is turned flat, parallel with the floor, and the neck is flexed such that a finger can be placed between the mandible and clavicle. Excessive rotation or flexion during positioning can cause neurovascular compromise, especially in patients with extracranial carotid disease or degenerative cervical spondylosis. Alternatively, the patient can be placed prone or in the lateral decubitus position. The skin incision starts above the auricle and curves behind the ear inferiorly 4 to 6 cm behind the mastoid and 6 to 8 cm behind the external auditory canal. Inferiorly, it extends just below the mastoid tip into the sternocleidomastoid muscle. If the incision is placed too far anteriorly, the scalp, muscle, and bone obscure visualization of the cerebellopontine angle. Surgical judgment should always prevail inasmuch as the surgeon should be familiar with surface skull landmarks that approximate the locations of the major sinuses. The asterion is an unreliable external landmark for the transverse-sigmoid junction, but a line drawn from the root of the zygoma to the inion (the superior nuchal line) is a good approximation of the transverse sinus. Typically, the transverse-sigmoid junction is avoided by placing the burr hole below this line, 2 cm below the asterion, two thirds of it behind and a third of it in front of the occipitomastoid suture. After the burr hole is completed, a craniotomy can be fashioned behind the sigmoid sinus and below the transverse sinuses. The bone up to the transverse and sigmoid sinuses can be rongeured to expose their edges. Alternatively, the mastoid air cells can be drilled directly to expose the sigmoid and transverse sinuses and then the craniotomy can be performed. The bone over the sigmoid and transverse sinuses can be drilled, first with a cutting burr and then with a diamond burr, and a thin shell of inner cortical bone left. The remaining thin shell of bone can be removed safely with a Penfield dissector or curet. Alternatively, a small linear incision, angled in the direction of the jugular bulbar, can be made over the cerebellar hemisphere. After the cerebellum is relaxed, the dural opening is completed in a curvilinear fashion with its base on the transverse-sigmoid junction. Intradurally, the junction of the tentorium and petrous bone at the superior and lateral extents of the dural opening should be visualized. FarLateralApproach the far lateral or transcondylar approach has several modifications and variations. It basically involves a partial condylectomy, with or without resection of the lateral mass of C1. This approach allows the surgeon to achieve an anterolateral trajectory to the brainstem, and it eliminates the need to traverse contaminated mucosal structures through the transoral or transfacial route. First described by Heros26 and later modified by Spetzler and Grahm,27 this approach provides access to lesions of the vertebrobasilar junction, inferolateral pons, anterolateral medulla, and upper cervical spinal cord. Its potential disadvantages include craniocervical instability, vertebral artery injury, wound infection, meningitis, or lower cranial nerve deficits. Various positions, including the modified park bench, sitting, lateral decubitus, supine with the head rotated, and lateral or half-lateral decubitus, have been used with this approach. At our institution, we prefer a modified park bench position in which the operating table is extended 10 to 20 cm by placing a rigid plastic board under the mattress. The dependent arm is cradled in a padded sling between the table edge and the Mayfield head holder (Codman, Inc. The clivus is brought perpendicular to the floor by performing four maneuvers on the neck: (1) flexion in the anteroposterior plane until the chin is one fingerbreadth from the clavicle, (2) rotation 45 degrees contralateral to the side of the lesion, (3) lateral flexion 30 degrees down toward the opposite shoulder (also the floor), and (4) slight distraction to increase the interval between the foramen magnum and C1 so that the surgeon can look down the axis of the brainstem and work between the horizontally oriented cranial nerves. The ipsilateral shoulder should also be retracted inferiorly with cloth tape to allow greater freedom of movement with the microscope. The patient should be taped securely to the bed to permit frequent and extreme rotation. Multiple skin incisions have been described, including the hockey-stick, lazy S, and simple paramedian linear incision. The traditional hockey-stick skin incision begins in the midline at C2 or C3, proceeds superiorly, and curves anteriorly and laterally to the mastoid tip. Monopolar cauterization is used to develop a plane below the superficial muscle fascia. A small muscle cuff is cut and left attached to the superior nuchal line for reapproximation of the fascia and muscles at the end of the procedure. Muscle is then stripped off the laminae of C1 and C2 with a Penfield dissector or gauze sponge. The advantage of this incision over the paramedian incision is that the dissection proceeds in a medialto-lateral direction; hence, the vertebral artery is approached more safely. A lateral decubitus position is used with the contralateral arm cradled beyond the edge of the bed. The lateral aspect of the ipsilateral C1 posterior arch is further removed with a rongeur to the lateral aspect of the dura. The extradural vertebral artery, which lies on top of the sulcus arteriosus, can be followed into the foramen transversarium with a Woodson or dental instrument. The venous plexus surrounding the artery can be the source of profuse bleeding, which can be controlled with Nu-Knit gauze and bipolar coagulation. The foramen transversarium can be unroofed by sliding the footplate of a Kerrison rongeur into it. The C2 nerve root between C1 and C2 should be preserved to avoid occipital numbness. The occipitoatlantal membrane is dissected off the foramen magnum with a curved curet to prepare for the craniotomy. A lateral suboccipital craniotomy extending from the midline down to the foramen magnum and laterally to the retromastoid region is fashioned. The lateral foramen magnum is drilled laterally to include the posteromedial third to half of the occipital condyle. This maneuver can be performed safely by drilling within the center of the condyle (with a diamond burr) and leaving a thin eggshell of bone that can be removed with rongeurs. The condylar emissary vein, when entered, should be expected to produce heavy venous bleeding, but it can be controlled with bone wax, cotton pads, and Nu-Knit. This positioning can be checked preoperatively with frameless stereotactic guidance. The craniotomy should extend above and below the transverse sinus to expose the junction of the traverse sinus and torcular. This craniotomy permits greater retraction of the tentorium superiorly than possible with a pure suboccipital craniotomy. It can be performed safely by placing a single burr hole lateral to the superior sagittal sinus with a pneumatic drill, footplate, and drill bit. Before the sinus is crossed, the surgeon should reverse the footplate and irrigate through the craniotomy line to confirm that the plate is extradural. The dura is opened in an inverted V shape with the base on the edge of the transverse sinuses. Bridging veins from the superior aspect of the cerebellum that drain into the transverse sinus are coagulated and divided to permit downward retraction of the cerebellum. If these veins are not coagulated early in the procedure, severe venous bleeding can occur during retraction. They must be coagulated as close to the surface of the cerebellum as possible to leave a pedicle on the surface of the tentorium. If avulsed at the interface of the tentorium, coagulation is not only futile but may enlarge the hole in the sinus and cause disastrous bleeding. If this occurs, a piece of Nu-Knit larger than the defect should be patched over the hole. A cotton pad is then placed over the hemostatic agent and can be held in place with a retractor during the remainder of the procedure. At the which should not be seen during the exposure, is situated anterior and medial to the anterior third of the condyle. The foramen transversarium can be unroofed with a rongeur or diamond burr, and the vertebral artery can be mobilized. The vertebral artery can be retracted medially and the lateral mass of C1 can also be drilled. The two limbs of the C-shaped dural opening are placed over the upper cervical cord and cerebellum, respectively, to increase the anterolateral exposure.
References
- Filipovich AH, Weisdorf D, Pavletic S, Socie G, Wingard JR, Lee SJ, et al. National Institute of Health consensus development project on criteria for clinical trials in chronic graft-versus-host disease: I. Diagnosis and staging working group report. Biol Blood Marrow Transplant 2005;11(12): 945-56.
- Lisk DR, Pasteur W, Rhoades H, et al. Early presentation of hemispheric intracerebral hemorrhage: prediction of outcome and guidelines for treatment allocation. Neurology. 1994;44(1):133-139.
- Pooley AS, Ewen P, Sutton CJ. Does transcervical resection of the endometrium for menorrhagia really avoid hysterectomy? Life table analysis of a large series. J Am Assoc Gynecol Laparosc 1998; 5: 229-35.
- Namen AM, Ely EW, Tatter SB, et al. Predictors of successful extubation in neurosurgical patients. Am J Respir Crit Care Med 2001;163(3):658-664.