

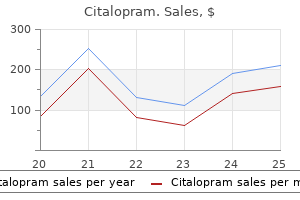
Citalopram
| Contato
Página Inicial
Stephen R. Thompson, MD, MEd, FRCSC
- Cooperating Associate Professor of Sports Medicine, University of Maine
- Medical Director, EMMC Sports Health
- Deputy Editor, The Journal of Bone and Joint Surgery
- Eastern Maine Medical Center Bangor, Maine
- Cofounder and Codirector, Miller Review Course Part II, Denver , Colorado
The genetic pathway was determined before the nature of the proteins had been analyzed medicine gif buy cheap citalopram 20 mg on line. Likewise brazilian keratin treatment buy citalopram paypal, gain-of-function alleles of let-23 act as if they are receiving a signal all the time treatment borderline personality disorder purchase 40 mg citalopram with mastercard, whether or not lin-3 is functional treatment low blood pressure citalopram 40 mg order overnight delivery, and thus activate the downstream signal-transduction cascade medicine 018 discount citalopram 20 mg mastercard, which in turn depends on having a functional allele of let-60. This pathway, called the epidermal growth factor signaling pathway, is conserved throughout animals, with inappropriate activation of the pathway leading to cancer. However, when the cell that would normally be a 1° cell is ablated, a cell that would normally have been a 2° cell differentiates into a 1° cell instead. It is thus unlikely that the absolute concentration of signal perceived is solely responsible for directing cell fate. This creates a feedback loop that reinforces the initial asymmetry between the 1° and 2° cells. Continued feedback between the signal and its perception amplifies the differences between the two cells, causing them to acquire distinct developmental fates. Cell Death During Development One of the striking observations made when Sulston, Brenner, and Horvitz tracked the fate of every cell during C. Of the 1090 cells produced during the development of a hermaphrodite worm, 131 cells undergo a process called programmed cell death, or apoptosis (introduced in Sections 3. This pathway is largely conserved across the animal kingdom (in humans, as well) and is a natural and important process that helps sculpt the development of tissues as well as maintain tissues in adult organisms. Indeed, it is estimated that 1011 cells are programmed to die every day in an adult human, many of them in epithelial tissues such as skin and intestine. Whereas loss-of-function mutants for genes in the apoptosis pathway are viable in C. Thus relatively simple animals such as Drosophila have a genome containing about 14,000 genes, whereas the human genome contains about 25,000 genes. Even organisms such as jellyfish and sea anemones possess genomes with gene numbers largely similar to those of vertebrates. Given this consistency of gene number, what is the biological explanation of how the presumed "complexity" of vertebrates is produced from a genetic toolkit that is similar to the one possessed by comparatively "simple" animals The answer seems to lie in the relative complexity of gene regulation rather than the invention of new genes for additional developmental processes. This proposal suggests that existing genes are recruited for new roles by means of changes in their regulation, both in space and time. Biologist Francois Jacob summed up this view of evolution when he said, "Evolution behaves like a tinkerer. It works on what already exists, either transforming a system to give it new functions or combining several systems to produce a more elaborate one. In this section, we consider an example of the co-option of genes by evolutionary "tinkering" to form newly evolved structures: digits (fingers and toes) on tetrapod limb appendages such as hands and feet. Evolution through Co-option Limb positioning in tetrapods (four-legged vertebrates) results in large measure from the expression of Hox genes that direct the anteriorposterior organization of the body. Work on chickens and mice demonstrates that expression of Hox genes along the anteriorposterior body axis defines the position at which a limb will develop. The expression of these two genes specifies the thoracic region of vertebrates, which is characterized by the formation of ribs from the vertebral column. Once limb positions are specified, cells of the mesenchyme (loosely connected sub-ectodermal cells) send a signal to the overlying ectodermal cells. The Sonic hedgehog (Shh) gene is orthologous to the Drosophila segment polarity gene hedgehog. Sonic hedgehog is expressed principally in the neural tube, where it helps organize the brain, eyes, and other structures through patterning of a group of cells known as the floor plate, and in developing limbs, where it directs the development of digits. The Case Study in this chapter discusses the consequences of different Shh mutations on mammalian development and morphology. All extant tetrapods are characterized by five or fewer digits in each set, and each digit in the set has a unique identity. If you allow your arms to hang straight down, you will see that your thumb (digit 1) is in the anterior position on your hand, while your pinky (digit 5) is in the posterior position. A second role of Shh in limb patterning is in the specification of digit identity. The Hox genes that play a conserved role in patterning the anteriorposterior axis in animals were considered candidates to be the genes acting downstream of Shh to specify the patterning events in digits. Despite the difference in position of hindlimb and forelimb along the body axis, the same five Hox genes are expressed in the developing digits of each limb. Their expression in the limb bud follows a precise temporal and spatial pattern and is dependent on Shh activity. The first gene to be expressed is Hoxd9, followed by Hoxd10, then Hoxd11, and so on through Hoxd13. Spatially, all genes share the same posterior boundary, but the anterior boundary of expression is different for each gene. Consequently, the five Hoxd genes subdivide the limb bud into five zones, each specified by a different combination of Hoxd gene expression. Analogous to patterning along the anteriorposterior axis, ectopic expression of different Hoxd genes within the developing limb bud results in transformations of digit identity. A similar combinatorial code of Hox gene expression also appears to specify the proximaldistal patterning of the limb buds themselves. Mutations that expand or increase Shh expression result in extra digits and have been documented in mice, chickens, dogs, cats, and humans. Finally, it is worth noting that the separation of the human limb bud into individual digits requires programmed cell death (see Section 18. These programs have been further modified during evolution in the secondary loss of legs in snakes and cetaceans. The loss of the front legs of snakes is due to an anterior shift in both Hoxc6 and Hoxc8 gene expression all the way to the base of the head. All vertebrae behind the snake head, except the first one, develop as thoracic vertebrae with ribs. In contrast, the convergent evolution of loss of hind legs in snakes and cetaceans is due to independent alterations in Shh activity in the developing hind limb bud. Hoxd expression during anteriorposterior patterning of the body axis, the changes would not result in defects of this earlier process. The acquisition of gene expression in the developing limb could be thought of as a gain-of-function mutation. The modularity of enhancers and silencers facilitates evolution by co-option because individual enhancer modules are free to evolve independently. Thus the patterning of a novel tetrapod organ, the limb, involved the co-option of, or tinkering with, preexisting genetic programs that already had developmental roles elsewhere. As noted above, a major constraint on this type of evolutionary change is that the more ancestral functions of the gene must not be disrupted. The two lineages of multicellular organisms you are likely to be most familiar with are animals and land plants. Since the common ancestor of plants and animals was a single-celled organism, multicellularity evolved independently in each lineage. Due to their independent origins, animals and plants differ in certain crucial aspects of their development. One difference is that germ-line cells in animals separate from somatic (body) cells much earlier in development than do the germ-line cells in land plants. Another difference is that animal cells are often motile during development, whereas plant cells are encased in a cell wall that essentially fixes them in the location at which they arise. Animals and land plants also differ with respect to when the basic form of the body plan takes shape. The animal body plan is established during embryogenesis, and subsequent development consists primarily of growth in size but without the addition of new organs. In contrast, throughout their lifetimes plants add new organs that are produced from pluripotent stem-cell populations. Finally, because plants often grow in a fixed location and are unable to migrate as many animals can, a plant must be able to alter its developmental program in response to changing environmental conditions throughout its lifetime. Thus, although identical twins in animals are nearly indistinguishable, genotypically identical plants may develop to look very different depending upon their growth environment. Despite these differences, developmental processes occurring in plants are remarkably similar to those in animals, especially in their reliance on the coordinated action of transcription factors and signaling molecules. Constraints on Co-option the ancestral roles of Hoxd genes pertained to patterning along the anteriorposterior axis of the body. Therefore, the role of Hoxd genes in specifying digit identity represents a co-option of function of already existing genes. These same ancestral genes also acquired roles in the differentiation of the nervous system floor plate, whose presence in all vertebrates is an indication that it evolved before limbs during vertebrate evolution. Limbs developed later within the tetrapod lineage, and in the course of limb evolution, Shh was co-opted to pattern digits, structures that did not previously exist. In the case of limb evolution, genes of the Hoxd cluster could have come under control of limb-specific enhancer modules leading to expression of the Hoxd genes in developing limbs. As long as changes in regulation did not disrupt Development at Meristems Plant development occurs at organized groups of pluripotent cells called meristems. The above ground parts of a plant are produced by shoot meristems and the below ground parts by root meristems. Meristems are generally indeterminate- that is, they can remain active for years, or in some cases the entire life of the plant. For example, the shoot meristem at the top of a pine tree can be active for centuries, continually producing leaves and side branches. Over time, the sizes of the central and peripheral domains remain remarkably constant. It is the continual production of new organs from meristems throughout the life of a plant that allows plants to adjust and adapt to changing local environmental conditions. The identity of the meristem determines what types of organs are produced from its periphery. Early in the life of a flowering plant, leaves are produced from the flanks of the shoot meristem, and roots are produced from the root meristem. At the upper side of the attachment point of the leaf to the stem an axillary meristem is formed, from which a branch can arise. This reiterative formation of meristems that produce leaves that produce branches containing meristems forms the basis of most aboveground development of flowering plants. In response to appropriate environmental conditions, the identity of meristems can change. For example, shoot meristems, which have been producing leaves, are converted in response to seasonal changes into reproductive meristems. A reproductive meristem may either develop directly into a flower meristem, or alternatively into an inflorescence meristem that produces flower meristems-an inflorescence being a group of flowers. Unlike the other meristems, flower meristems are determinate: no more stem cells are available after the flower meristem has produced a fixed number of organs. Because each type of meristem is characterized by a specific pattern of gene expression, mutations in key genes can result in homeotic transformations of meristem types. Combinatorial Homeotic Activity in Floral-Organ Identity Several flowering plant species have been adopted as models for the study of genetics. For example, peas (Pisum sativum), with which Mendel performed his experiments, and maize (Zea mays), in which transposons were discovered, were introduced in earlier chapters. Since the 1980s, study of homeotic mutants in Arabidopsis and another plant species, Antirrhinum (snapdragon), has led to insights into the genetic basis of flower development and revealed developmental parallels with animals. The outermost whorl is occupied by sepals, organs that protect the flower bud during development. The second whorl is occupied by petals, which in many species attract pollinators. Stamens, the male organs that produce pollen, are located in the third whorl, and the female organs-carpels, containing the ovules- occupy the central whorl. A second class, the B-class mutants, exhibit homeotic transformations in the middle two whorls, where sepals replace petals and carpels replace stamens, so that the four whorls consist of sepals, sepals, carpels, and carpels. In C-class mutants, homeotic transformations in the third and fourth whorls result in flowers where petals develop in the positions normally occupied by stamens, and the cells that would normally give rise to the carpels behave as if they were another flower meristem that reiterates the developmental cycle. Similar mutants can be found in a number of ornamental plant species and are often referred to as "double flowers. Additive double-mutant phenotypes suggest that the two genes do not interact, whereas nonadditive double-mutant phenotypes suggest that the two genes interact to influence a common developmental pathway. For example, in apetala2 agamous flowers, the first and fourth whorls have leaf-like carpels whereas the second and third whorls are occupied by organs with features of both petals and stamens. The agamous mutation has a phenotypic effect in the first and second whorls in an apetala2 background (compare the identities of these whorls in an apetala2 single mutant with a apetala2 agamous double mutant), an effect not observed in a wild-type background, where phenotypic defects of agamous are limited to the third and fourth whorls. Thus, each whorl is characterized by a different combination of homeotic gene activity that specifies floral organ identity. The A-class activity by itself in the first whorl specifies sepals, A@class + B@class in the second whorl specifies petals, B@class + C@class in the third whorl specifies stamens, and C-class by itself in the fourth whorl specifies carpels. To account for the mutant phenotypes (specifically the apetala2 agamous mutant described above), a second postulate of the model is that the A-class and C-class activities are mutually antagonistic, so that in an A-class mutant background, C-class activity is found in all four whorls; and conversely, in a C-class mutant background, A-class activity is in all four whorls. The specification of identity by combinations of homeotic gene activities and cross-regulatory interactions between the floral homeotic genes is reminiscent of specification of segmental identity in Drosophila by Hox genes. These observations suggest that since floral organs are evolutionarily derived from leaves, one role of the floral homeotic genes is to modify a leaf into a specialized floral organ. Would reverse or forward genetics approaches be more suited to identifying the genes required for early kelp development
Pneumolysin binds to the Fc portion of immunoglobulins and activates the classical pathway medicine 018 purchase citalopram without prescription. It has been suggested that cell wall components symptoms xanax addiction 20 mg citalopram order overnight delivery, pneumolysin medicine jokes discount citalopram online mastercard, and other proteins such as PepO are released by the bacteria to deplete complement (168 medicine 4h2 pill effective citalopram 20 mg, 172 treatment integrity purchase citalopram with paypal, 173). This would have the most direct impact during blood stream infections and in the lungs. LytA is responsible for pneumococcal lysis in the stationary phase as well as in the presence of antibiotics (174). Furthermore, it has been shown that LytA is important for the release of capsule in response to antimicrobial peptides found on the epithelial surface (78). Autolysin-mediated lysis is responsible for the spike in inflammation observed immediately following antibiotic treatment of meningitis. Cytotoxicity In addition to the pathology derived from inflammation, the pneumococcus can directly damage eukaryotic cells. All teichoicated species contain ChoP, a key component increasing inflammatory activity. These mediators combine to produce the symptomatology of pneumococcal infection, including changes in blood flow, fluid balance in the tissue, and leukocytosis. Studies using a variety of challenge routes and animal models have convincingly demonstrated that pneumolysin-deficient mutants are drastically attenuated (38, 92, 96, 177). The mechanism responsible for pneumolysin secretion from the bacteria is poorly understood. Pneumolysin has three functionally separate domains: one activating complement, one causing hemolysis, and the other binding to cholesterol. Biofilm formation enhances fomite survival of Streptococcus pneumoniae and Streptococcus pyogenes. Regev-Yochay G, Raz M, Dagan R, Porat N, Shainberg B, Pinco E, Keller N, Rubinstein E. Nasopharyngeal carriage of Streptococcus pneumoniae by adults and children in community and family settings. Molecular surveillance on Streptococcus pneumoniae carriage in non-elderly adults; little evidence for pneumococcal circulation independent from the reservoir in children. Molecular surveillance of nasopharyngeal carriage of Streptococcus pneumoniae in children vaccinated with conjugated polysaccharide pneumococcal vaccines. The epidemiology of invasive pneumococcal disease in Alaska, 1986-1990: ethnic differences and opportunities for prevention. Otitis media in young Aboriginal children from remote communities in Northern and Central Australia: a cross-sectional survey. Epidemiology of nasopharyngeal carriage of respiratory bacterial pathogens in children and adults: cross-sectional surveys in a population with high rates of pneumococcal disease. Age-specific cluster of cases of serotype 1 Streptococcus pneumoniae carriage in remote indigenous communities in Australia. The effects of season and age on pneumococcal acquisition and carriage in the first 24 months of life. Influence of child care on only released as a result of autolysis (178), it has also been shown to have an independent route, because mutants lacking the autolytic enzyme, LytA, show the same pattern of release as the wild-type bacteria (179, 180). Pneumolysin is a pore-forming toxin that kills cells via necroptosis, a programmed mode of necrosis. At high concentrations, pneumolysin, like other pore-forming toxins, triggers necroptosis due to ion dysregulation (181, 182). The toxin binds to cholesterol on the surface of the host cell and oligomerizes to form pores as large as 30 nm in diameter (183). At lower concentrations, the toxin has a variety of effects on different cell types. Pneumolysin has been demonstrated to slow ciliary beating of epithelial cells (184), disrupt tight junctions (185), and inhibit the capacity of neutrophils and macrophages to kill by inhibiting oxidative burst (186, 187). Disruption of the alveoli-capillary barrier contributes to the leakage that allows serous exudates to enter the lungs and the bacteria to cross into the blood stream (188). In the middle ear, pneumolysin is responsible for damage to the cochlea and hair cells, contributing to hearing loss (72). During meningitis, pneumolysin causes neuronal damage mediated by an influx of extracellular calcium triggering apoptosis (146). Hydrogen peroxide (H2O2) is a major product of pneumococcal metabolism and damages host tissues. Mutation of spxB dramatically attenuates virulence in the respiratory tract but not in the blood stream (38). Studies of the cytotoxic effects of H2O2 are not as comprehensive as those for pneumolysin. Nonetheless, H2O2 also contributes to mitochondrial damage of neurons, resulting in apoptosis (146), and inhibits beating of ciliated ependymal cells lining the ventricular system of the brain and cerebral aqueducts (189, 190). Asymptomatic colonization allows the pneumococcus to persist in the population, and extensive serotype diversity complicates the development of effective vaccines. Streptococcus pneumoniae: Invasion and Inflammation nasopharyngeal carriage of Streptococcus pneumoniae and Haemophilus influenzae. Acquisition and invasiveness of different serotypes of Streptococcus pneumoniae in young children. Age- and serogroup-related differences in observed durations of nasopharyngeal carriage of penicillinresistant pneumococci. Impact of pneumococcal conjugate vaccines on nasopharyngeal carriage and invasive disease among unvaccinated people: review of evidence on indirect effects. Trends in drug resistance, serotypes, and molecular types of Streptococcus pneumoniae colonizing preschool-age children attending day care centers in Lisbon, Portugal: a summary of 4 years of annual surveillance. The changing epidemiology of invasive pneumococcal disease in aboriginal and non-aboriginal western Australians from 1997 through 2007 and emergence of nonvaccine serotypes. Pneumococcal bacteremia: no change in mortality in 30 years: analysis of 104 cases and review of the literature. Polysaccharide capsule composition of pneumococcal serotype 19A subtypes is unaltered among subtypes and independent of the nutritional environment. Effectiveness of pneumococcal polysaccharide vaccine for preschool-age children with chronic disease. Effects of pneumococcal conjugate vaccine 2 years after its introduction, the Netherlands. Effect of pneumococcal conjugate vaccination on serotype-specific carriage and invasive disease in England: a cross-sectional study. Association of intrastrain phase variation in quantity of capsular polysaccharide and teichoic acid with the virulence of Streptococcus pneumoniae. Relationship between cell surface carbohydrates and intrastrain variation on opsonophagocytosis of Streptococcus pneumoniae. A novel solenoid fold in the cell wall anchoring domain of the pneumococcal virulence factor LytA. Identification of the teichoic acid phosphorylcholine esterase in Streptococcus pneumoniae. The molecular characterization of the first autolytic lysozyme of Streptococcus pneumoniae reveals evolutionary mobile domains. Defense mechanisms involving Fc-dependent functions of immunoglobulin A and their subversion by bacterial immunoglobulin A proteases. Rosenblut A, Napolitano C, Pereira A, Moreno C, Kolhe D, Lepetic A, Ortega-Barria E. Etiology of acute otitis media and serotype distribution of Streptococcus pneumoniae and Haemophilus influenzae in Chilean children <5 years of age. Viral-bacterial interactions and risk of acute otitis media complicating upper respiratory tract infection. Acute otitis media in pediatric medicine: current issues in epidemiology, diagnosis, and management. Cloning and characterization of nanB, a second Streptococcus pneumoniae neuraminidase gene, and purification of the NanB enzyme from recombinant Escherichia coli. Evaluation of the virulence of a Streptococcus pneumoniae neuraminidase-deficient mutant in nasopharyngeal colonization and development of otitis media in the chinchilla model. Comparison of alteration of cell surface carbohydrates of the chinchilla tubotympanum and colonial opacity phenotype of Streptococcus pneumoniae during experimental pneumococcal otitis media with or without an antecedent influenza A virus infection. A role for pneumolysin but not neuraminidase in the hearing loss and cochlear damage induced by experimental pneumococcal meningitis in guinea pigs. Cytotoxic effects on hair cells of guinea pig cochlea produced by pneumolysin, the thiol activated toxin of Streptococcus pneumoniae. The human polymeric immunoglobulin receptor binds to Streptococcus pneumoniae via domains 3 and 4. Streptococcus pneumoniae contains 3 rlrA pilus variants that are clonally related. Expression of the Streptococcus pneumoniae pilus-1 undergoes on and off switching during colonization in mice. Nontypeable pneumococci can be divided into multiple cps types, including one type expressing the novel gene pspK. PspK of Streptococcus pneumoniae increases adherence to epithelial cells and enhances nasopharyngeal colonization. The streptococcal lipoprotein rotamase A (SlrA) is a functional peptidyl-prolyl isomerase involved in pneumococcal colonization. Surfaceassociated lipoprotein PpmA of Streptococcus pneumoniae is involved in colonization in a strain-specific manner. High levels of genetic recombination during nasopharyngeal carriage and biofilm formation in Streptococcus pneumoniae. Streptococcus pneumoniae biofilm formation and dispersion during colonization and disease. Interkingdom signaling induces Streptococcus pneumoniae biofilm dispersion and transition from asymptomatic colonization to disease. Dynamic changes in the Streptococcus pneumoniae transcriptome during transition from biofilm formation to invasive disease upon influenza A virus infection. Epidemiology of otitis media during the first seven years of life in children in greater Boston: a prospective, cohort study. Causative pathogens, antibiotic resistance and therapeutic considerations in acute otitis media. Effect of neuraminidase on the expression of the 3-fucosyl-N-acetyllactosamine antigen in human tissues. Deacetylation of sialic acid by esterases potentiates pneumococcal neuraminidase activity for mucin utilization, colonization and virulence. Role of neuraminidase in lethal synergism between influenza virus and Streptococcus pneumoniae. Synergistic stimulation of type I interferons during influenza virus coinfection promotes Streptococcus pneumoniae colonization in mice. Effect of antiviral treatment on the outcome of secondary bacterial pneumonia after influenza. Streptococcus pneumoniae endopeptidase O (PepO) is a multifunctional plasminogen- and fibronectin-binding protein, facilitating evasion of innate immunity and invasion of host cells. Jensch I, Gámez G, Rothe M, Ebert S, Fulde M, Somplatzki D, Bergmann S, Petruschka L, Rohde M, Nau R, Hammerschmidt S. PavA of Streptococcus pneumoniae modulates adherence, invasion, and meningeal inflammation. The effects of pneumolysin and hydrogen peroxide, alone and in combination, on human ciliated epithelium in vitro. Induction of pulmonary inflammation by components of the pneumococcal cell surface. Streptococcus pneumoniae anchor to activated human cells by the receptor for plateletactivating factor. B-arrestin 1 participates in platelet-activating factor receptor-mediated endocytosis of Streptococcus pneumoniae. The phosphorylcholine epitope undergoes phase variation on a 43-kilodalton protein in Pseudomonas aeruginosa and on pili of Neisseria meningitidis and Neisseria gonorrhoeae. Decoration of lipopolysaccharide with phosphorylcholine: a phasevariable characteristic of Haemophilus influenzae. Bacterial peptidoglycan traverses the placenta to induce fetal neuroproliferation and aberrant postnatal behavior. Pilus adhesin RrgA interacts with complement receptor 3, thereby affecting macrophage function and systemic pneumococcal disease. Purified Streptococcus pneumoniae endopeptidase O (PepO) enhances particle uptake by macrophages in a Toll-like receptor 2- and miR155-dependent manner. Dissecting bacterial cell wall entry and signaling in eukaryotic cells: an actindependent pathway parallels platelet-activating factor receptor-mediated endocytosis. Severe pneumococcal pneumonia causes acute cardiac toxicity and subsequent cardiac remodeling. Platelet-activating factor receptor and innate immunity: uptake of Gram-positive bacterial cell wall into host cells and cell-specific pathophysiology. Circulating pneumolysin is a potent inducer of cardiac injury during pneumococcal infection. Cell invasion and pyruvate oxidase derived H2O2 are critical for Streptococcus pneumoniae mediated cardiomyocyte killing. Rifampin followed by ceftriaxone for experimental meningitis decreases lipoteichoic acid concentrations in cerebrospinal fluid and reduces neuronal damage in comparison to ceftriaxone alone.
Order 20 mg citalopram otc. Private STD Check Fonda village NY Best Private STD Check NY.
Sequence requirements for target activity in sitespecific recombination mediated by the Int protein of transposon Tn1545 medicine 5113 v proven citalopram 40 mg. The mef(E)-carrying genetic element (mega) of Streptococcus pneumoniae: insertion sites and association with other genetic elements treatment 0f osteoporosis purchase generic citalopram canada. Variation on a theme; an overview of the Tn916/Tn1545 family of mobile genetic elements in the oral and nasopharyngeal streptococci treatment effect citalopram 20 mg line. Composite mobile genetic elements disseminating macrolide resistance in Streptococcus pneumoniae symptoms chlamydia generic citalopram 40 mg with amex. Nucleotide sequence analysis of the termini and chromosomal locus involved in site-specific integration of the streptococcal conjugative transposon Tn5252 897 treatment plant rd discount citalopram 40 mg buy on line. Characterization of a genetic element carrying the macrolide efflux gene mef(A) in Streptococcus pneumoniae. Del Grosso M, Iannelli F, Messina C, Santagati M, Petrosillo N, Stefani S, Pozzi G, Pantosti A. Macrolide efflux genes mef(A) and mef(E) are carried by different genetic elements in Streptococcus pneumoniae. Nucleotide sequence and functional map of pC194, a plasmid that specifies inducible chloramphenicol resistance. Analysis of Streptococcus pneumoniae sequences cloned into Escherichia coli: effect of promoter strength and transcription terminators. Interruption of capsule production in Streptococcus pneumonia serotype 3 by insertion of transposon Tn916. Method for introducing specific and unmarked mutations into the chromosome of Streptococcus pneumoniae. An rpsL cassette, janus, for gene replacement through negative selection in Streptococcus pneumoniae. A modified Janus cassette (Sweet Janus) to improve allelic replacement efficiency by high-stringency negative selection in Streptococcus pneumoniae. Genome evolution driven by host adaptations results in a more virulent and antimicrobial-resistant Streptococcus pneumoniae serotype 14. A high incidence of prophage carriage among natural isolates of Streptococcus pneumoniae. Comparative genomic analysis of ten Streptococcus pneumoniae temperate bacteriophages. The Escherichia coli MutL protein physically interacts with MutH and stimulates the MutH-associated endonuclease activity. Homeologous recombination and mismatch repair during transformation in Streptococcus pneumoniae: saturation of the Hex mismatch repair system. Hyperrecombination in Streptococcus pneumoniae depends on an atypical mutY homologue. The recA gene of Streptococcus pneumoniae is part of a competence-induced operon and controls lysogenic induction. Dominant role of nucleotide substitution in the diversification of serotype 3 pneumococci over decades and during a single infection. Genome analysis of a highly virulent serotype 1 strain of Streptococcus pneumoniae from West Africa. Donner J, Bunk B, Schober I, Spröer C, Bergmann S, Jarek M, Overmann J, Wagner-Döbler I. Genome evolution to penicillin resistance in serotype 3 Streptococcus pneumoniae by capsular switching. Expression of the lux genes in Streptococcus pneumoniae modulates pilus expression and virulence. Pneumococci are the largest cause of community-acquired pneumonia in the developed world. The rate of meningitis in children in the United States is about 4 cases per 100,000 children, with a fatality rate of about 15% (2). In the developing world pneumococci are an important cause of childhood deaths due to bacterial respiratory infection following viral disease. Recently, about one-third to one-half of pneumococci recovered from humans in the United States have been found to be at least partially resistant to penicillin, and penicillinresistant strains are frequently also resistant to other common antibiotics (3). The rise of antibiotic resistance among pneumococci has already complicated treatment, especially of meningitis (4), and threatens to greatly increase the morbidity and mortality caused by pneumococci unless new control measures are developed. It has long been recognized that the best management of most infectious disease is prevention. Vaccines offer the prospect of a highly cost-effective means of preventing morbidity and mortality caused by pneumococci. This article provides a concise summary of issues critical to the development and application of pneumococcal vaccines. There are several relatively recent reviews that address this topic in more detail (511). In the preantibiotic era, vaccination attempts utilized whole killed pneumococci injected parenterally. Although such vaccines were sometimes protective in humans, they were also highly reactogenic. These killed vaccines were mainly used to elicit antibody in animals for passive treatment of infected humans (12). Only about one-third of the greater than 90 serotypes occur with a significant frequency in adult infections, and the distribution of common serotypes is even more restricted in children (6, 1720). However, during this time antibiotics were becoming widely available, and these drugs diminished the perceived impact of pneumococcal infections. The two commercially available pneumococcal vaccines were withdrawn from production for lack of demand (7). Despite a growing selection of antibiotics and increasingly sophisticated critical care technology, morbidity and mortality from invasive pneumococcal infections have remained high. Antibiotics and supportive care alone are not sufficient to completely eliminate the impact of pneumococcal disease because in many cases deaths occur so quickly that antibiotics are not effective (21). A significant impediment to conducting the most rigorous controlled clinical trials has been the ethical concern of denying at-risk groups an approved and recommended vaccine. The ability to make antibodies to protein antigens appears quite rapidly after birth, and T cell-dependent antibody production is highly efficient. This strategy has proven highly successful in preventing both invasive disease and colonization with Haemophilus influenzae type b (33). A large clinical trial in infants using a four-dose schedule of immunization with a heptavalent conjugate vaccine proved this strategy to be highly effective in preventing invasive (bacteremic) disease (30, 36, 37). However, this conjugate vaccine has shown somewhat more modest efficacy in preventing pneumonia (15) and otitis media (38). Since the vaccine serotypes that colonize and infect children include most of the serotypes associated with antibiotic resistance, and since children carry pneumococci at a much higher frequency than adults, there are expectations that childhood immunization may reduce the transmission of antibiotic-resistant pneumococci in the entire population (3942). It has been observed, however, that significant replacement carriage and otitis media occur with capsular types not present in the vaccine (38, 39, 42). Since the introduction of the 7-valent vaccine, a significant increase in invasive disease was seen among nonvaccine strains, which led to the introduction of the 13-valent conjugate vaccine in 2010 (4345). However, despite the introduction of the new conjugate vaccine, strain replacement still persisted in carriage and invasive disease, with rates of invasive disease due to nonvaccine strains ranging from 57. Also of concern are observations that immunization with the conjugate vaccine alters the bacterial flora of the upper respiratory tract in children and has affected the pathogens recovered from children with carriage and acute otitis media (51, 52). A potentially worrisome trend has been noted in increased rates of carriage of Staphylococcus aureus in children immunized with the pneumococcal conjugate vaccine (51, 53). The distribution of pneumococcal serotypes commonly causing invasive disease in adults is much larger than that for children. This recommendation was based on the results of a large efficacy trial in the Netherlands, which showed 75% and 46% efficacy against vaccine type invasive disease and pneumococcal pneumonia, respectively, among people over 65 years (55). The 13-valent conjugate vaccine is also recommended for routine use among people over the age of 19 with immunocompromising conditions, functional or anatomic asplenia, cochlear implants, or cerebrospinal fluid leak (54, 58). Attempts are also underway to develop conjugate vaccines with much higher valence to broaden coverage of different serotypes (59, 60). An approach with multiple antigens that collectively protects against carriage, pneumonia, and sepsis, delivered through a mucosal route, would be expected to be optimally protective. PspA and PspC Pneumococcal surface protein A (PspA) is produced by all pneumococci (70). PspA also appears to reduce phagocytosis of pneumococci even in the absence of C and antibody (75). PspA may also play a role on mucosal surfaces since it blocks killing by apolactoferrin (76). Antibody to PspA reverses this blockage and enhances killing by apolactoferrin (76). Although serologically variable when examined with monoclonal antibody, PspA is highly cross-reactive when examined with polyclonal sera (70, 77). Monoclonal and polyclonal antibodies to PspA can passively protect mice from otherwise fatal bacteremia and otherwise fatal sepsis caused by pneumococci (78, 79). Parenteral immunization with PspA has been able to protect mice against fatal infections, and this protection has been observed to be highly cross-protective regardless of PspA type (7981). Intranasal and transdermal immunization with PspA has also been shown to protect against nasopharyngeal carriage in an adult mouse carriage model, raising the possibility that vaccines could be developed to prevent carriage and transmission of pneumococci in addition to invasive disease with pneumococci (45, 61, 8286). A group in Japan has used PspA for the development of a mucosal vaccination protocol which does not use an immunogenic adjuvant or molecules that can follow the nerves from the nose into the brain. Their protocol and the transdermal route could be particularly important in moving PspA and other mucosal immunogens into humans as the vaccines routes able to elicit both mucosal and systemic immune responses (83, 84). A number of pneumococcal proteins, including PspA, PspC, and LytA, are able to bind choline and share similar, and sometimes indistinguishable, choline-binding domains near the C-terminal ends of the proteins (8790). The N-terminal end of PspA is composed of a largely coiled-coil a-helical sequence (89, 91) that is responsible for most of the cross-protective immunity elicited by PspA (92, 93). Paradoxically, this region also contains significant structural and serologic variability within the PspA molecule (92, 94). In spite of its variability at the amino acid sequence level, PspAs are very cross-reactive (70, 79) and can be divided into families based on their cross-reactivity and amino acid sequences (77, 94). Ninety five percent or more of clinical isolates belong to families 1 and 2 (77, 94, 95). Immunization of humans with a single recombinant family 1 PspA leads to antibodies that are able to protect mice from otherwise fatal infection with strains expressing either family 1 or family 2 PspAs (80). However, other data indicate that the best vaccine would probably contain PspAs from both family 1 and family 2 (81, 93, 96). In addition, it has been shown that the 80-120 amino acid long proline-rich domain in the middle of PspA, is also able to elicit protective antibody (97). The use of well-chosen alpha-helical and proline-rich domains in immunizing PspA molecules should ensure coverage of diverse PspAs. The serum of virtually all adults, and most children over 7 months of age contains detectable antibody to PspA (98, 99). The levels of antibody are higher in adults than in children, and it is thus possible that natural antibodies to PspA will contribute more to the immunity to pneumococcal infection in adults than in young children. However, serotype coverage remains limited, and it is unlikely that significantly more than 13 to 15 of the 98 currently known serotypes will be included in future conjugate formulations. Data from the original conjugate vaccine trials indicated that, although carriage of vaccine types was reduced, the vacated niche was promptly occupied by nonvaccine serotypes known to cause invasive disease in humans (8). Such "replacement" carriage, as well as replacement disease due to nonvaccine serotypes, has occurred to varying extents in virtually all regions where conjugate vaccines have been widely used since the licensure of the original 7-valent formulation in 2000 and continued after introduction of 10- and 13-valent formulations (6567). In addition, the cost of the conjugate vaccines remains very high; thus, without philanthropic support, their use in developing countries, where the need for effective pediatric vaccines is the greatest, will remain restricted. In view of this, much attention has focused on the possibility of developing vaccines based on pneumococcal protein antigens common to all serotypes (5, 68, 69). Such proteins, being T cell-dependent antigens, are likely to be highly immunogenic in human infants and able to elicit immunological memory. The vaccine potential of various pneumococcal protein antigens are discussed further below. Pneumococcal Vaccines 365 immunization to elicit high levels of antibodies to PspA will be able to enhance protection of young children and adults with waning immunity from infections with pneumococci. PspC is a protein that has some similarity to PspA in its proline-rich and choline-binding domains (100). The gene for this protein was originally identified through its close similarity to PspA (101). However, the alpha-helical domain of PspC is more complex than that of PspA and is present as several very distinct alleles with distinct combinations of functions (102, 103). This protein has been independently discovered by others based on its ability to bind secretory IgA (SpsA) (104), choline (CbpA) (105), and factor H (Hic) (106). It has potential roles in colonization, adherence, and invasion (103105, 107, 108). Immunity to PspC is able to protect against pneumococcal infection and carriage (103, 108). Pneumolysin All pneumococci produce pneumolysin, a potent 53-kDa thiol-activated pore-forming cytolysin. This cell-free pneumolysin can attack any cell that has cholesterol in its plasma membrane (111). In addition to its cytotoxic properties, cell-free pneumolysin is capable of directly activating the classical complement pathway away from the pneumococcal surface and does so in the absence of specific antibody. Classical pathway activation by pneumolysin reduces the local concentration of reactive C3 and other classical pathway complement components.
These observations suggest that similar to other Gram-positive bacteria symptoms 0f ms purchase 20 mg citalopram otc, sortasedependent display of proteins occurs in S symptoms 4dpiui buy citalopram 20 mg cheap. The pneumococcal adr gene encoding the O-acetyltransferase has been initially identified in a mariner mutagenesis screen for increased sensitivity to penicillin in the background of the resistant strain Pen6 (65) medicine while breastfeeding generic citalopram 40 mg free shipping. Adr mutant cells also show higher sensitivity to lysozyme symptoms stiff neck purchase citalopram uk, and Adr shares sequence homology with the peptidoglycan O-acetyltransferase OatA of S symptoms yeast infection discount citalopram 20 mg amex. Recent work showed that peptidoglycan O-acetylation by Adr occurs at midcell and that this modification protects the cell wall from cleavage by the autolysin LytA (67). Sortase A-Dependent Covalent Attachment of Proteins to the Pneumococcal Cell Wall Most virulence-related pneumococcal proteins studied so far were shown to be attached to the cell surface through the choline residues of teichoic acids, and pneumococci have been described as a paradigm for the display of virulence proteins through specific but noncovalent associations with the cell surface (68). Such choline-binding proteins include PspA, PsaA, the enzyme phosphoryl choline esterase (69, 70), and LytA, in which a 20-amino-acid repeat was shown to recognize choline residues in wall or membrane teichoic acids (71). Analysis of the penicillin-resistant South African clinical isolates revealed that they produced cell walls of a radically different composition from the one seen in the penicillinsusceptible and nonencapsulated laboratory isolate (75). The same determinants were also described independently and named fib by another group (76). The Cell Wall of Streptococcus pneumoniae 289 branches to the epsilon amino group of lysine residues in the staphylococcal peptidoglycan (77). Inactivation of the murM operon or inactivation of the murM gene alone did not interfere with growth of the bacteria but caused the production of a peptidoglycan composed exclusively of linear muropeptides. Another consequence of inactivation was the complete loss of the penicillin-resistant phenotype. Analysis of the murM alleles from penicillin-resistant isolates showed that they differed from one another and from the murM carried by penicillin-susceptible strains in regions of considerable sequence diversity that were distributed as heterologous "patches" along the murM gene sequence. All transformants remained penicillin susceptible, but their cell wall composition changed in directions which corresponded to the muropeptide pattern of the strain from which the murM allele was derived. The relationship between murM alleles and the penicillinresistant phenotype is less clear. Interestingly, in this particular experiment the penicillin-resistant secondary transformants began to show "fitness" defects: defective growth and premature autolysis in antibiotic-free medium, similar to the defective physiology observed in laboratory isolates of penicillin-resistant pneumococci (81). In these early studies, a structural relationship between C-polysaccharide and lipocarbohydrate was suggested, and in contrast to the various strain-specific capsular polysaccharides, lipocarbohydrate and C-polysaccharide were considered pneumococcal common antigens. This was confirmed by serological methods which showed that all 90 known capsular types of S. In 1980, Jennings and coworkers published the first complete structure of pneumococcal teichoic acid (11). The functionally important phosphocholine residues are phosphodiester-linked to O6 of the N-acetyl-Dgalactosaminyl residues. Species with one phosphocholine per repeat are distinguished from species with two by higher mobility of the individual bands. Both types of teichoic acid have identical chains (top) which carry phosphoryl choline and D-alanine residues. The figure was kindly provided by Nicolas Gisch (Research Centre Borstel, Germany). Many teichoic acid genes cluster in three genomic regions, called lic1, lic2, and lic3. The adjacent lic2 region contains the tacF gene, encoding the putative teichoic acid flippase, and the licD1/licD2 genes, encoding phosphotransferases for decoration of the teichoic acid subunits with phosphoryl choline residues. The lic3 region locates elsewhere on the chromosome and was identified by the presence of a gene, called licD3, with sequence similarity to licD1 and licD2, which is surrounded by four other putative teichoic acid genes. LicD3 is most likely the phosphotransferase for the transfer of ribitol phosphate during synthesis of the repeating unit, which involves eight additional enzymes that synthesize the repeating unit linked to the carrier lipid bactoprenol phosphate. Presumably, Spr1222 polymerizes the teichoic acid precursors to the nascent chains, which are still linked to the carrier lipid, before these are flipped across the cytoplasmic membrane by TacF. Interestingly, TacF is specific for choline-loaded teichoic acid precursor chains, ensuring that the mature teichoic acids contain phosphoryl choline residues (94). Interestingly, mutants lacking tacL grow normally under laboratory conditions but show attenuated virulence in mouse models of acute and/or systemic infections by S. This modification introduces positive charges to teichoic acids and thus increases resistance to cationic antimicrobial peptides. Pce carries an N-terminal signal sequence and contains a C-terminal choline-binding domain with 10 homologous repeating units similar to those found in other pneumococcal surface proteins. The catalytic (phosphorylcholine esterase) activity is localized on the N-terminal part of the protein. The product of the enzymatic digestion of 3H-choline-labelled cell walls was shown to be phosphorylcholine. Sequence comparison indicates that Pce, the protein originally identified through its unique enzymatic activity (99), is identical to choline-binding protein E (CbpE), described by Masure and colleagues as a protein implicated in the attachment of pneumococci to nasopharyngeal cells (100). The crystal structure of Pce shows a catalytic site with two zinc ions and an elongated binding domain recognizing the phosphocholine residues of teichoic acid (101). There are several proteins in pneumococci-including LytA, LytB, and LytC-that specifically recognize and bind to phosphocholine residues. These proteins contain distinct domains which are responsible for their specific biological activities, whereas the choline-binding domains are homologous and contain 6 to 10 choline-recognizing repeats of 20 amino acids each. The structure of crystallized LytA and LytC and their interaction with choline residues in the cell wall have been described (107, 108). However, the requirement of LytA for built-in phosphocholine residues is no longer seen when solubilized cell wall polymers are degraded to muropeptides (12) or when the teichoic acid is removed from peptidoglycan (48). Only the phosphate content of cell walls was reduced, consistent with the absence of phosphocholine. In vivo, the lack of active autolysins became apparent by impaired cell separation at the end of cell division and by resistance against stationary phase and penicillin-induced lysis. In the case of strain R6Chi and related strains, the lack of a nutritional requirement for choline was explained by the loss of specificity of the teichoic acid flippase TacF for choline-loaded teichoic acid precursor chains (94). R6Chi contains a single point mutation in the tacF gene, rendering the cells choline independent. When growing in the presence of choline, R6Chi still incorporated phosphocholine into its teichoic acids, suggesting that the mutated TacF can transport choline-loaded and -unloaded teichoic acid precursor chains. Pneumococci grown in ethanolamine-containing medium show several striking abnormalities: unlike the cholinegrown bacteria, pneumococci utilizing ethanolamine grow in long chains and are completely resistant to the cell walldegrading activity of LytA (103). Upon addition of trace amounts of radiolabeled choline to a culture grown on ethanolamine, the bacteria immediately shifted to the utilization of choline so that the nascent wall units that began to incorporate into the cell surface contained choline residues in the teichoic acid component of the nascent cell wall and produced regions that were susceptible to hydrolysis by exogenous LytA enzyme added to the medium. It was possible to show by electron microscopy that under these conditions the LytA enzyme performed an enzymatic "microsurgery" on the bacteria: it has selectively removed a thin equatorially located band of cell wall, thus identifying the anatomical site of wall incorporation and growth zone (58). Another abnormality of the ethanolamine-grown pneumococci, the complete inhibition of cell separation, has allowed the design of experiments to test the mode of inheritance of pneumococcal cell walls. Pneumococci labeled in their wall by titrated choline were shifted to an ethanolaminecontaining medium in which the bacteria continued to grow in the form of chains of cells, i. Since the teichoic acid choline does not exhibit turnover during growth, the localization of radioactively labeled cells within the chains of bacteria could provide clues as to the mode of wall segregation. The finding was that the radioactive label remained in large clusters in association with cells that were located either at the tips or at the center of chains. The results demonstrate the conservation of large hemispherical segments of the cell wall which are passed on intact to daughter cells during cell division (112). Novel methods such as staining nascent peptidoglycan with fluorescent vancomycin (113), incorporating fluorescent D-amino acids (114), and superresolution microscopy (32) confirmed that pneumococci incorporate new cell wall units into the preexisting wall material at a single growth zone located at midcell. Cell Wall Growth and Cell Division Complexes the mode of growth and cell wall segregation described above suggests that S. Nevertheless, the characteristic ovococcal shape of pneumococci suggests that there must be major differences in the cell wall growth mechanism compared to those of the rods. One central difference is that pneumococci, despite conservation of all the other components of the elongation complex, lack the rod-shape determinant MreB, i. The second difference is that, despite previous reports (2), both components of the elongasome and divisome show a clear septal localization, with no substantial variation in timing of recruitment, although they may show differences in localization profile. Subsequent studies supported this model and provided insights into the coordination and regulation of the complex during the cell cycle (120123). However, it was not possible to assess directly the effect of inactivation of these genes until genetic systems allowing the generation of merodiploids and conditional lethal mutants became available. Moreover, recent work employing a clustered regularly interspaced short palindromic repeat interference method (41) provides an additional powerful tool to confirm results obtained with other methods and identify and characterize new essential genes. However, some of these genes were not essential in strain R6, a laboratory derivative of D39, and similar strains (53, 122124). Presumably, these laboratory strains contain preexisting suppressor mutations that could compensate for the loss of otherwise essential genes. This result is also supported by the fact that pbp2a, which shows a synthetic lethal relationship with pbp1a, can be readily inactivated in strain R6 (44). A previously uncharacterized gene, spd 0768 (spr0777 in R6), encoding the membrane protein named CozE (for coordinator of zonal elongation) was identified in a Tn-seq screen for genes dispensable in D39 with inactivated pbp1a (118). With the exception of pcsB and gpsB, which showed strain-dependent essentiality, inactivation of cell division genes was more consistent between strains. Photoactivated localization microscopy showed that as in other bacteria, the Z-ring in S. Following the fluorescent tagged-FtsZ localization and the Z-ring diameter, it was observed that FtsZ molecules assemble at midcell at the beginning of the cell cycle to form a single patchy ring, which thickens and then disassembles during constriction, while new Z-rings assemble at the future division sites (or new equators) of the newborn cells. Notably, unconstricted double Z-rings, suggesting short-lived intermediates, were observed in a small percentage of cells at midcell but not at the future division site (127). Once the Z-ring is formed, the later pneumococcal cell division proteins localize to midcell, and a few to midcell and cell poles, in exponentially growing pneumococcal cells. A hierarchical order of recruitment to the septum has not been determined, and it may not even be a linear sequence of events, although colocalization studies based on fluorescence microscopy suggest that divisome assembly occurs in at least two steps. In the absence of MreB, it is still unknown how elongation components are targeted to midcell and how lateral and septal growth are coordinated. FtsZ is believed to be the best candidate to act as a scaffold to coordinate both side-wall and septal synthesis (127), although this role has not been experimentally verified, and the effect on S. A recent study characterized conditional lethal mutants for the cell division protein FtsA, providing insights about how integration of the elongation and septation machineries at a single mid-cell location in S. Unexpectedly, the complete depletion of FtsA in the pneumococcus resulted in cell ballooning and, ultimately, lysis, in sharp contrast to the cell filamentation phenotype observed in rods (121). FtsZ rings and peptidoglycan synthesis delocalized upon FtsA inactivation, suggesting that the cells could neither elongate nor divide. In contrast, inactivation of genes encoding other cell division components, such as GpsB (5) and SepF (128), which in B. Consistent with these results, and in contrast to the model rods where FtsA localizes after FtsZ, in S. Taken together, these observations led to the hypothesis that in oval-shaped cocci, the actin-like FtsA, and not FtsZ, could play a major role in coordinating peripheral and septal peptidoglycan synthesis, likely carrying out a role similar to the actin-like MreB in preseptal synthesis in E. However, since FtsA is needed to tether FtsZ to the membrane (131) and FtsZ may be required for FtsA to be targeted to the septum, additional work is necessary to distinguish any direct role for FtsA in coordinating both modes of peptidoglycan synthesis from that of FtsZ. This approach identified the membrane-bound, likely endolytic transglycosylase, MltG (see above), which is involved in cell elongation (53). Two independent studies also identified Spr1851/ Spd 1849, either as a novel substrate for phosphorylation by the Ser/Thr protein kinase, StkP, in the laboratory strain S. Experiments with null or phosphoablative (T89A) alleles of eloR in the R704 strain revealed that both pbp2b and rodA could be deleted, while its phosphomimetic form (T89D or T89E) could only be tolerated in strains that acquired suppressor mutations in mreC and rodZ (133). In contrast, no phenotype was observed in either KhpB phosphoablative or phosphomimetic derivatives in S. Despite discrepancies between strains, these studies highlight the crucial yet complex role of StkP in the regulation of the pneumococcal cell cycle. Regulators of Cell Wall Growth and Division and Role of Phosphorylation Similar to other Gram-positive bacteria, S. Several StkP substrates playing a role in cell wall metabolism and cell division were identified to be phosphorylated in vivo in a global study of the pneumococcal phosphoproteome (139). However, only some of them have been confirmed to be specifically phosphorylated by StkP in vitro and/or in vivo. Inactivation of stkP, or overproduction of PhpP, resulted in elongated cells with multiple and often unconstricted division rings, perturbed in cell wall synthesis. These data indicate that StkP and PhpP play an important role in coordinating cell wall synthesis during growth and division to achieve and maintain the characteristic ovococcal shape. StkP was proposed to act as a molecular switch that, through phosphorylation of key division substrates, signals the shift from peripheral to septal cell wall synthesis (143). A parallel study of the role of StkP in pneumococcal cell division partially confirmed these results and proposed diverse functions for the different StkP domains in the R6-derived strain R800 (142). The round and chaining phenotype, connected with the stkP deletion or truncation (142), was not observed in other studies and was interpreted to be likely due to differences in genetic background, growth conditions, or suppressor mutations in specific stkP null strains (39). R800 cells deleted for gpsB were viable but displayed an elongated and twisted-towel phenotype, more severe than that previously reported for S. The Cell Wall of Streptococcus pneumoniae 297 moniae D39 cells depleted of gpsB, where this gene is essential (5). Both FtsZ and peptidoglycan synthesis were shown to have a helical pattern of localization in DgpsB R800 cells, in which phosphorylation of all StkP substrates, including itself, was abolished, consistent with the StkP delocalization observed. In this model, GpsB, which has been shown to be phosphorylated in Streptococcus agalactiae (149) and B. Intriguingly, this study revealed that the mreD mutant differed from the expected morphology observed in other studies for mreD mutants in the same or similar genetic backgrounds (39, 118, 124), highlighting once more the difficulties in understanding individual effects of gene inactivation in S. The CbpD-dependent lysis data were then correlated with protein-protein interactions between the identified proteins using the bacterial two-hybrid analysis system (152). The regulatory function in cell wall biosynthesis of its paralog, GpsB, appears to become more clear.
References
- Rau R, Hockel S. [Piroxicam gel versus diclofenac gel in active gonarthroses]. Fortschr Med 1989; 107(22):485-8.
- Charghi A, Charbonneau J, Gauthier G: Colocystoplasty for bladder enlargement and bladder substitution: a study of late results in 31 cases, J Urol 97:849, 1967.
- Hagenbeek A, Zijlstra JM, Lugtenburg P, et al. Transplant BRaVE: combining brentuximab vedotin with DHAP as salvage treatment in relapsed/refractory hodgkin lymphoma. A phase 1 dose-escalation study. Haematologica 2016;101(s5):44.
- Tawk RG, Villalobos HJ, Levey EI, Hopkins LN. Surgical decompression and coil removal for the recovery of vision after coiling and proximal occlusion of a clinoidal segment aneurysm: technical case report. Neurosurgery. 2006;58(6):E1217; discussion E1217.
- Chang GJ: Open and laparoscopic surgery for colon cancer. In Pollock RE, Curley SA, Ross MI, Perrier ND (eds) Advanced Therapy in Surgical Oncology. BC Decker Inc, Hamilton, 2008, pp 266-280.
- Budoff MMJ. Cardiac CT Imaging: Diagnosis of Cardiovascular Disease. Berlin: Springer; 2006.