

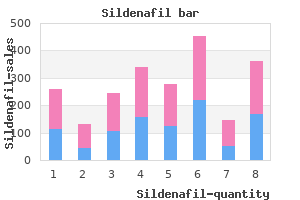
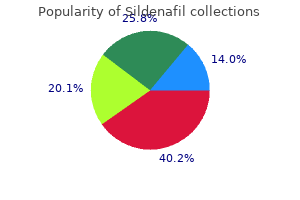
Sildenafil
| Contato
Página Inicial
R. Bryan Bell, MD, DDS, FACS
- Clinical Associate Professor
- Oregon Health and Science University
- Attending Head and Neck Surgeon and Director of
- Resident Education
- Oral and Maxillofacial Surgery Service
- Legacy Emanuel Medical Center
- Portland, Oregon
Glycogen synthase exists as two major isoforms and is regulated by both allosteric and covalent modification [12] acupuncture protocol erectile dysfunction sildenafil 25 mg visa. Glycogen synthase activity is generally suppressed by phosphorylation erectile dysfunction statistics age purchase discount sildenafil line, especially at site 2 (ser 7) [12] xyzal impotence purchase 75 mg sildenafil. Glycogen synthase is potently activated by allosteric interaction with glucose-6-phosphate [80] erectile dysfunction doctors in connecticut buy discount sildenafil 100 mg. Phosphorylated glycogen synthase (deactivated) requires regulatory phosphatases for reactivation impotence at 40 100 mg sildenafil order with mastercard. Phosphatase activity is organized by scaffolding proteins known as glycogen targeting subunits [81]. In the previous section of this chapter, we discussed distinct enzymes that have evolved to circumvent the otherwise irreversible glycolytic steps and allow gluconeogenesis to occur. Thus, the forward reactions of glycolysis serve to produce pyruvate and also moderate gluconeogenesis during feeding. However, despite the elegance of the regulatory mechanisms of glycolysis, liver is, first and foremost, a gluconeogenic organ and gluconeogenesis remains modestly active even during the unfavorable conditions of a glucose load [42]. This residual activity provides a mechanism to prevent lactate build-up, and is the basis for indirect glycogen synthesis. Glycogen metabolism as a therapeutic target for diabetes One approach to reducing hepatic glucose production during diabetes is to improve glycogen storage by activation of glycogen synthase and/or inhibition of glycogen phosphorylase. However, glycogen synthase kinase also phosphorylates many other targets, making its mechanism difficult to decipher. These compounds improve glycemia in animal models of diabetes by increasing direct and indirect glycogen synthesis [83]. The adverse effects of pharmacologic promotion of glycogen storage are not completely known, but long term glycogen phosphorylase inhibition in animal models of type 2 diabetes can cause hepatomegaly, glycogenosis, and steatosis, side effects also common to glycogen storage diseases. Glycolysis Glucose-6-phosphate not used for direct glycogen synthesis can be metabolized in the glycolytic pathway. About half of the glucose taken up by liver during duodenal glucose absorption will undergo glycolysis to pyruvate. The resulting pyruvate can be used for lipogenesis and amino acid synthesis, lactate production, or cycled back to glucose-6-phosphate for indirect glycogen synthesis. Seven are catalyzed by enzymes with equilibrium constants that allow the reaction to proceed in either the "forward" (glycolytic) or "reverse" (gluconeogenic) direction dependent upon physiologic changes in the relative concentrations of substrates and products of the reactions. Three enzymatic steps, glucose phosphorylation by glucokinase (see earlier), the conversion of fructose-6-phosphate to fructose-1,6-bisphosphate by phosphofructokinase, and the conversion of phosphoenolpyruvate to pyruvate by pyruvate kinase are considered essentially irreversible due to a large release of free energy. Biochemical and molecular studies revealed that the synthesis and degradation of F-2,6-P2 is catalyzed by a single protein with two distinct catalytic sites [87]. This protein was named 6-phosphofructose-2-kinase/fructose-2,6-bisphosphatase but is also known more simply as "the bifunctional enzyme. Phosphorylation at this site inhibits the 2-kinase activity of the enzyme, and increases its bisphosphatase activity, resulting in a net decrease in F-2,6-P2 concentration within cells. Regulation of glucose metabolism in liver 207 In the transition from the fasted to the fed state, this regulation is reversed. Following a mixed meal, insulin and glucose levels rise in the blood while glucagon is decreased. These conditions cause dephosphorylation of the bifunctional enzyme, restoring its 2-kinase activity and causing a rapid increase in F-2,6-P2 levels. This represents a "feed-forward" mechanism by which flux through an early step in glycolysis activates a later step. Summary the liver has been studied for its metabolic characteristics for more than 200 years. During fasting, liver releases glucose from glycogen stores and uses gluconeogenesis to produce glucose from pyruvate, lactate, amino acids, and glycerol. During prolonged fasting, gluconeogenesis is upregulated to compensate depleted glycogen levels and prevent major changes in blood glucose concentration. Upon feeding, liver rapidly converts to an organ that utilizes more glucose than it produces. Postprandial glucose uptake by liver replenishes glycogen and provides substrate for lipid synthesis for long-term energy storage. A large proportion of glycogen is formed by "indirect" synthesis, a process driven by the simultaneous activity of glycolysis and gluconeogenesis. These elegant shifts in metabolism are regulated by transcription, posttranslational modification, allosteric and the substrate regulation of several key enzymes in the gluconeogenic and glycolytic pathway. Glucagon and other counterregulatory hormones activate transcription factors and co-activators that induce the expression and stability of gluconeogenic enzymes, while insulin opposes these effects and stimulates glucose uptake. American Journal of Physiology-Endocrinology and Metabolism 1979;237(3):E214Â223. American Journal of Physiology-Endocrinology and Metabolism 2001;281(3):E413Â419. Cheng Z, Guo S, Copps K, et al: Foxo1 integrates insulin signaling with mitochondrial function in the liver. Flux control coefficients of the enzymes in the gluconeogenic pathway in the absence and presence of glucagon. American Journal of Physiology-Endocrinology and Metabolism 1993;265(4): E636ÂE647. American Journal of Physiology-Endocrinology and Metabolism 2007;292(2):E400Â407. To a much lesser extent than in plants, glucose also constitutes a building block for structural and enzymatic components of cells as well as the extracellular matrix. As a metabolic substrate, glucose is present in organisms essentially in its simple, monomeric form (-D-glucopyranose) and as a branched polymer of -glucose, namely glycogen. Disaccharides of glucose (lactose, maltose, and sucrose) are quantitatively less important. Glucose is present in plasma water at a concentration that-in a healthy adult who has fasted overnight-ranges between 3. A family of proteins residing in the plasma membrane (and in microsomal membranes) can specifically and reversibly bind glucose molecules, and transfer them across cell membranes in both directions. They differ from one another in both tissue distribution and physiologic regulation, particularly with respect to sensitivity to insulin stimulation. It is usual for more than one species of glucose transporter to be expressed in a tissue. Variable dominance of the other types of glucose transporters is found in tissues in which glucose metabolism does not respond to insulin acutely (erythrocyte, liver, kidney, brain, pancreatic cells). The rate of glycolytic utilization of transported glucose by the erythrocyte mass is estimated to be 25 mol min-1 or about 6 mol Ч min-1 Ч m-2 of diffusion surface. Since this rate is 17,000 times less than the rate of glucose transport in these cells, glucose concentration will be the same in plasma and erythrocyte water. Similarly, the total amount of glucose leaving a body region is the product of whole-blood glucose level in the venous effluent times the blood flow rate. Thus, under steady-state conditions of blood flow (F), arterial glycemia (A) and organ metabolism, the net balance of glucose movement across a body region is given by the product of blood flow and the arteriovenous (A-V) whole-blood glucose concentration International Textbook of Diabetes Mellitus, Fourth Edition. It is evident that the use of plasma flow rate and plasma glucose concentration systematically underestimates the net organ balance of glucose (and, for that matter, of any substance that travels in plasma as well as in erythrocytes, for example lactate and some amino acids). In fact, plasma flow is less than blood flow by an amount equal to the hematocrit (40%), while plasma glucose is higher than whole-blood glucose by only 15% (0. This is the structural basis for the long-held notion that brain and placental glucose utilization are not regulated by insulin, so that vital functions for the adult and fetal organism can be maintained in the face of variable metabolic conditions. In general, diffusion of glucose from the intravascular compartment into the interstitial fluid space is very rapid. Whether this is accomplished by facilitated diffusion mediated by specific cell transporters (as in the brain) or by simple diffusion through intercellular clefts of endothelial cells is still uncertain. A lower limit of glucose diffusion through endothelial membranes is represented by cellular glucose uptake (which does not include glucose back-diffusion into the intravascular compartment), and can be estimated by carrying out the following calculation. Thus, about 200 billion glucose molecules must pass through each capillary surface each minute to travel through the interstitial space. Direct measurement of interstitial glucose concentrations has proven to be difficult, and has yielded conflicting results. Difficult as it may be to measure or calculate this "bathing" concentration of glucose, it is this concentration that dictates the activity of cellular glucose transport together with the state of specific activation of the glucose transporter. In addition to substrate mass action, cellular glucose uptake is influenced by changes in blood flow and hormonal stimulation by insulin. Thus, glucose metabolism is regulated by a distributed control between tissue glucose delivery (blood flow), transit through the interstitium, plasma membrane glucose transport and glucose phosphorylation (hexokinase) [6,7]. Baron [9] was amongst the first to show that physiologic hyperinsulinemia, while maintaining euglycemia, stimulated muscle blood flow resulting in enhanced muscle glucose uptake. The insulin-mediated increase in muscle blood flow results from two separate effects: (i) relaxation of resistance of vessels, and (ii) recruitment of previously unperfused muscle tissue secondary to relaxation of terminal arterioles [8]. This recruitment effect of insulin has been elegantly documented using microbubbles in combination with contrast-enhanced ultrasonography in rodents and humans [8]. The result is expansion of the capillary surface area for nutrient and insulin delivery. Impaired insulin-mediated vasodilation is associated with metabolic resistance to the stimulatory effect of insulin on muscle glucose utilization in type 2 diabetic and obese nondiabetic individuals [8,9]. Evidence in humans has demonstrated that the insulin concentration in lymph and interstitial fluid is considerably less than in plasma [10,11]. Transport rates of large molecules, such as insulin, into the interstitial space, and thus the cell membrane, is slow. Insulin binding to its receptor in the vascular endothelium and subsequent incorporation into caveolae are involved in the transcytosis of the hormone from the intravascular to the interstitial space [8]. Since the rate of insulin entry into skeletal muscle appears to be a critical step for insulin action, elucidation of the molecular and anatomical mechanisms involved in the transvascular process is likely to produce novel insights into the etiology of metabolic insulin resistance. Thus, in the only organs in the body in which the presence of glucose-6-phosphatase (G6Pase)-the enzyme catalyzing the formation of free intracellular glucose from glucose-6-phosphate (G6P)-makes glucose available to the circulation, the transporter is of a type that only responds to the concentration gradient between the internal and external side of the plasma membrane. This ensures that, when insulin is around to stimulate inward glucose transport in tissues with sensitive transporters, the liver can release glucose into the blood stream as long as sufficient G6P is derived from glycogenolysis or gluconeogenesis (or both) and sufficient G6Pase activity is there to accumulate free glucose on the inside of the plasma membrane. Under these conditions of reversed gradient, the presence of insulin-responsive glucose transport Insulin actions in vivo: glucose metabolism 213 activity on hepatocyte cell membranes would enhance glucose outflow, thereby opposing the plasma glucose-lowering action of insulin. In contrast, physiologic control of the direction and rate of glucose flux through the hepatocyte membrane is not on the transport step but on intracellular processes. In summary, the differential distribution and acute insulin sensitivity of the various classes of glucose transporters provide the backbone for the functional characteristics of glucose diffusion and exchange in the body. On the whole, free glucose is present in blood water, interstitial fluid, and the intracellular water compartment of insulin-independent tissues (liver, brain, kidney, intestine, placenta) in total amounts which, in the overnight-fasted healthy adult, average 80 mmol (14 g or 1. Free glucose is found at concentrations that (a) are uniform in the intravascular water compartment; (b) decline across the interstitial space toward the cell; (c) fall precipitously within cells that consume glucose avidly. The regional characteristics of tissue composition, blood flow rate, capillary density. Glycogen is present in most cells in cytoplasmic granules that encase the enzymes that regulate its metabolism. In normal humans, the largest part of glycogen stores is in liver and skeletal muscle. In the former, 3Â4 g of glycogen are packed in each 100 g of parenchyma; in striated muscle, the concentration is much lower (0. Thus, approximately 25 times more glycosyl units are stored in intracellular depots as glycogen than are dissolved in body water as free glucose. Glycogen metabolism is controlled by irreversible cascades of enzymatic reactions ultimately acting upon the proximal enzymes that catalyze glycogen synthesis (glycogen synthase) and degradation (glycogen phosphorylase) (see subsequent discussion). Key points · Glucose concentration is on average 15% higher in the plasma than the whole-blood glucose under ordinary circumstances. The extent of these differences depends on the circulatory region and on its rate of glucose utilization. Glucose metabolism Introduction Any given concentration of glucose in plasma (and in the space in equilibrium with plasma) is the result of simultaneous release of glucose into the circulation and uptake of glucose from the blood stream into cells. Whenever plasma glucose concentration is stable, the concurrent rates of its release and overall uptake must be equal. Accepted terminology 214 Chapter 14 refers to stationary conditions of glucose concentrations and flux rate as the "steady state," with the understanding that any physiologic steady state is only an approximation of the true, ideal steady state. Furthermore, the "glucose system" refers to the whole space or volume into which glucose is present as free glucose, regardless of how many compartments this system consists of, where they are physically located in the body, and how they are interconnected. Finally, the reference pool for glucose kinetics customarily is the plasma (or whole blood), for in most clinical or experimental circumstances the plasma is the only site accessible for sampling. When the plasma glucose concentration changes over time, one rate of glucose flux (entry or removal) is being exceeded by the other. Under these non-steady-state conditions, the glucose rates of entry and removal are conventionally termed rate of appearance (Ra) and disappearance (Rd), respectively. The glucose system is strongly homeostatic with respect to glucose levels, in that the normal variations in human plasma glucose concentration throughout a day of life are confined within a surprisingly narrow range. It should be recalled that glucosuria is called upon whenever glycemia exceeds the renal threshold, 180 mg dL-1, as if a safety measure had been set to cope with emergency when metabolic control fails. These considerations alone indicate that the body does not tolerate either hypoglycemia or hyperglycemia. For the former, the obligate dependence of brain function on the use of glucose as fuel classically has been offered as a rational explanation. For hyperglycemia, the evidence-and hence the concept-that high glucose levels, if not immediately life-threatening, are nonetheless intolerable to bodily functions, is more recent but no less compelling, and currently goes under the name of "glucose toxicity" (see Chapter 27). Methods Under conditions of an overnight fast, the liver accounts for 80% of glucose production with the remaining 20% coming from the kidney [12,13]. Placing catheters across the splanchnic bed (one in a hepatic vein and another in any artery) and measuring splanchnic blood flow. However, the liver and extrahepatic splanchnic tissues (gut, pancreas, spleen, etc.
This delay can describe the sustained insulin secretion response often observed at the end of the test erectile dysfunction treatment machine cheap sildenafil 75 mg buy on-line. The second approach assumes that the sustained response is due to potentiation of insulin secretion [19 erectile dysfunction under 40 cheap sildenafil generic,20] erectile dysfunction natural cures order sildenafil 100 mg amex. The latter approach appears to have more experimental support erectile dysfunction protocol foods order sildenafil 50 mg with visa, as potentiation phenomena have been clearly demonstrated in vivo in man erectile dysfunction from adderall discount sildenafil online visa. Several tests have been developed for this purpose and this section describes the most relevant and the characteristics of insulin secretion that they reveal. The hyperglycemic clamp the most typical test for the study of -cell response employs a brisk and sustained elevation of glucose concentration from a baseline value. The -cell response to this glucose stimulus is biphasic, with an initial insulin secretion burst lasting about 5Â8 min (first-phase secretion), followed by a drop towards basal levels and then by a relatively rapid increase that persists as far as hyperglycemia is maintained (second-phase secretion). One important physiologic feature highlighted by the hyperglycemic clamp is that the cell responds to the increase in glucose in a proportionate manner. This dose-response can be assessed with greater accuracy using a stepped increase in glucose concentration, the so-called graded glucose infusion test [22]. In this test, glucose is infused intravenously at increasing rates and glucose and C-peptide are measured. Insulin secretion is calculated by deconvolution of C-peptide and the -cell dose-response is constructed by plotting insulin secretion rates at the end of each glucose infusion period versus the corresponding glucose concentration. The -cell dose-response is a fundamental characteristic of insulin secretion; its slope, which represents the sensitivity of the cell to glucose, is a key -cell function parameter. In healthy subjects insulin secretion at a basal glucose level of 5 mmol L-1 is about 90 pmol min-1 and increases by five- to Normal -cell function 111 (a) 5 mmol LÂ1 1000 800 600 400 200 0 Â40 Â20 0 9 mmol LÂ1 Glucose concentration [24]. Impairment of first-phase secretion is already present in subjects at risk of developing diabetes [25], and is predictive of diabetes onset [26]. Because of these relevant characteristics, assessment of first-phase secretion has been widely used. This widely used index is typically calculated as the mean increment above baseline in insulin concentration in the first 8Â10 min. Moreover, similarly to the hyperglycemic clamp, parameters of insulin sensitivity can be derived as well [28]. Many of the studies concerning the relationships between insulin sensitivity and insulin secretion, discussed later, are based on this approach [29]. In summary, insulin secretion when assessed with intravenous glucose challenges appears to be highly dynamic with two main characteristic phases, a rapid and transient one followed by a more sustained one. These responses can be reproduced in vitro, by pancreas perfusion as well as in perifused islets [9,32]. Nonetheless, the mechanisms underlying the biphasic response remain only partially understood. It has been proposed that first-phase secretion is the consequence of the discharge of a pool of insulin granules located in the proximity of the cell membrane in response to an increase in intracellular calcium triggered by a cascade of electrochemical events generated by glucose utilization inside the cell [33]. Exocytosis, however, is very sensitive to calcium levels and the typical early peaking of intracellular calcium [34] may well exert a direct contribution to this phenomenon. Second-phase secretion is also controlled by calcium, but glucose itself, independently from calcium, plays a role in sustained insulin secretion [35]. In the hyperglycemic clamp, glucose con- centration is briskly elevated from the basal level using a suitable intravenous glucose infusion (a). The sharp glucose increase elicits a rapid and short-lived insulin secretion peak (first-phase secretion), followed by a drop towards basal levels and then by a relatively rapid return to a sustained value in the second half of the clamp (second-phase secretion). The hyperglycemic clamp assesses insulin secretion at two steady-state glucose levels (fasting and elevated glucose); a rudimentary -cell dose-response can be determined from these data (b). The relationship is fairly linear within this glucose concentration range and above; under these circumstances, -cell glucose sensitivity. First-phase secretion becomes evident with a brisk elevation of glucose concentration. The amount of insulin secreted during the first phase is dependent on the magnitude of the glucose increase, so that even the first phase recognizes a dose-response feature [9]; in a typical +7 mmol L-1 hyperglycemic clamp it is around 4 nmol m-2 representing the 10Â15% of what is secreted per hour in the second phase [23]. Though limited, the amount of insulin secreted in the first phase is relevant at least for two reasons. First, while the secretion burst is apparent with a rapid elevation in glucose concentration, the underlying secretory mechanisms appear to be active even for a more gradual rise in glucose concentration [9]. These mechanisms are likely to be responsible for a response that is anticipated compared to what would be predicted solely on the basis of the dose-response and this anticipation has relevant physiologic implications for glucose homeostasis, as discussed later. The insulin response to glucose ingestion is more pronounced than that elicited by an intravenous infusion achieving the same glucose levels. Estimates of the magnitude of the potentiating effect of oral glucose in normal subjects vary from study to study, partly because of the different methods used to calculate the incretin effect. Recent studies based on modeling analysis report an increase in insulin secretion of 1. The incretin effect appears to be quite variable also within subjects with normal glucose tolerance, ranging from a negligible effect to a two- to threefold amplification. The incretin effect is dose-dependent, that is, higher glucose doses elicit stronger effects; for a fivefold increase in the glucose load, the increase in incretin effect is almost twofold, from 1. The action of these hormones is glucose-dependent, 550 500 Insulin secretion (pmol/min/m2) 450 400 350 300 250 200 150 100 50 8. At similar glucose levels, insulin secretion is considerably increased during oral glucose ingestion (b). Consequently, the -cell dose-response relating insulin secretion to glucose concentration is shifted upwards (c). Normal -cell function 113 Insulin secretion that is, the increase in insulin secretion is higher when glucose concentration is higher, whereas at low glucose the amplification is marginal. The relationship between the levels of incretin hormones and the incretin effect are in fact generally weak [38,47,48]; whether this is due to limited assay precision or factors other than the hormone levels remains to be clarified. Therefore, based on this analysis the contribution of early secretion phenomena is quantitatively limited during an oral glucose load as well. The response to oral glucose is also characterized by phenomena that enhance -cell glucose sensitivity during the test, referred to as "potentiation. The -cell dose-response explains most of the insulin secretion excursions (bottom, gray line). However, in the initial period insulin secretion is relatively higher (bottom, horizontally banded area); this anticipative response accounts for 10Â15% of the total suprabasal secretion. However, some studies have reported that the -cell dose-response does not remain the same during the 24 hours. In particular, in relation to the glucose levels insulin secretion is relatively potentiated during the morning meal and attenuated during the following meals and the night, to a significant extent [20,50]. After the ingestion of a small amount of proteins (30Â50 g) [51,55] or a larger amount of proteins (2 g kg-1) [56], plasma insulin was raised two- to threefold over baseline and remained persistently elevated for 90 or 240 minutes, respectively. In support of this hypothesis, it has been shown that insulin response to different protein solutions was closely related to the increase of plasma amino acids [57Â59] with large differences among the individual amino acids unexplained by the functional group [60]: phenylalanine and glycine shown to be the most potent (+30 pmol L-1), histidine, tyrosine, and, surprisingly, arginine as neutral, and the others as intermediate (+10. In vitro studies [61] have reported that the insulinotropic effect of amino acids is mostly dependent on the amino acid type, duration of exposure and concentration; furthermore only combination of amino acids stimulates insulin secretion when added at physiologic concentrations, whereas higher concentrations of individual amino acids are required to activate insulin secretion [61]. Accordingly, the intravenous administration of a relatively small number of amino acids individually promotes insulin release while the maximum stimulus is elicited by mixed amino acids [62]. In contrast to other amino acids, homocysteine in vitro showed a dose-dependent negative effect on insulin secretion in pancreatic cells [63]. When either amino acids or proteins were ingested with glucose, plasma insulin levels were not significant or only slightly different from those after oral glucose alone but at lower glucose concentration [52Â55,59,60,64,65] suggesting a direct effect on -cell glucose sensitivity. A 24Â48-h lipid infusion has been reported to reduce [78], increase [79] or not significantly change [80] glucose-induced insulin secretion. Slow -cell response modes and adaptation mechanisms the secretion mechanisms discussed above are relatively rapid and typically suited to coping with the insulin needs of a meal. The healthy cell can also respond with slower modes, adapting to insulin demand if required by the metabolic conditions. Glucose-induced potentiation of insulin secretion Slow response modes are unveiled by prolonged exposure to hyperglycemia or repeated glycemic stimuli. During a hyperglycemic clamp, particularly at high glucose levels (>10 mmol L-1), the second-phase response exhibits a slow progressive rise over time [23]. When two consecutive hyperglycemic episodes are brought about, the insulin response after the second one is higher compared to the first [11]. With a prolonged (three day) infusion of glucose at a low constant rate, the insulin response assessed with a hyperglycemic clamp before and after the infusion is increased more than twofold [93]. Similarly, a prolonged glucose infusion makes the -cell dose-response, assessed with the graded glucose infusion test, steeper [22]. When glucose is infused intravenously to mimic the response to an oral glucose test, increasing glucose doses and concentrations produces an upward shift of the -cell dose-response, that is, insulin secretion becomes greater for the same glucose level [38]. The common denominator of these phenomena is that sustained hyperglycemia potentiates insulin secretion; this mechanism provides an additional resource to control glucose levels. Normal -cell function 115 Another classical experiment showing the potentiating effect of exposure to hyperglycemia involves the use of arginine as a secretagogue. When the arginine bolus is administered in a hyperglycemic state created by a hyperglycemic clamp, the insulin secretion response is potentiated compared to the basal state [94]. In normal subjects, the magnitude of this potentiation increases almost in proportion to the glucose levels until it reaches a plateau above 30 mmol L-1 glucose, where the response to arginine is more than fivefold the response at basal glucose. The initial slope of this curve, denoted as the glucose potentiation slope, is an index of the ability of glucose to potentiate insulin secretion [94]. Adaptation to insulin sensitivity A widely studied adaptive mechanism of the cell is the modulation of its response by prevalent insulin sensitivity. Thus, insulin-resistant subjects with normal glucose tolerance secrete more insulin to cope with the increased insulin demand. According to this paradigm, a more appropriate index of -cell function is the product of the insulin secretion and sensitivity indices, as on the hyperbola representing normal compensation this index is constant. The product of the insulin secretion and sensitivity indices has been historically denoted as the "disposition index" [28]. The disposition index, rather than the absolute insulin secretion response, should reflect intrinsic -cell function. This has been criticized because it has been shown that the assumption of a hyperbolic relationship is not valid for all indices [97,98]. In the presence of inverse relationships of other kinds, the disposition index provides a biased assessment of -cell function [98]. This implies that compensation for insulin resistance is of particular importance for the tuning of fasting insulin secretion and therefore fasting glucose, while the secretion mechanisms that regulate postprandial glucose are less influenced by this form of adaptation. The changes in glucose levels elicited by these hormones obviously will be detected by the cell, which will respond by enhancing insulin secretion. Yet, all these hormones, as well as others and the nervous system exert a coordinated direct effect on the cell resulting in an integrated and sophisticated control network. The typical example is the intra-islet networking encompassing the entangled interaction between, and cells [100]. Glucagon released from the cell has a stimulatory effect on insulin secretion by the cell, an effect that is commonly used to test residual endogenous insulin secretion in vivo [101]. The latter requires, however, pharmacologic doses of glucagon (1 mg), while the small increase in circulating insulin levels observed with glucagon infusion is more likely the result of the hyperglycemic effect of the hormone. Insulin secretion, in turn, exerts an inhibitory effect on glucagon secretion, whereas somatostatin, released from cells, suppresses secretion of both insulin and glucagon [102], and therefore, is believed to integrate pancreatic islet hormonal effect. More recently an interaction between somatostatin and ghrelin has been reported in regulating insulin release [103]. On top of the direct effect on the cell, resulting in suppression of both basal and stimulated insulin secretion, somatostatin may exert an effect on insulin secretion also through modulation of a number of gastrointestinal hormones, for instance by inhibiting, with different potency, the release of gastrointestinal hormones [104]. An even greater inhibitory effect has been claimed to be exerted by obestatin [106] though variable and divergent effects may be elicited by different levels of the hormone [107]. Claude Bernard first suggested the involvement of the nervous system in the regulation of insulin secretion. His hypothesis was then supported by the discovery by Langerhans that pancreatic islets were indeed highly innervated. Subsequently it was found that parasympathetic, sympathetic, and sensory nerve endings are present that can affect insulin and the -cell function through the release of a number of neurotransmitters [112]. The response of the cell to nerve stimulation is the result of the balance between the stimulatory effects of the parasympathetic nerve endings and the inhibitory one of the sympathetic nerves. However, much of the information is derived from animal studies, whereas investigation into the innervation of the human islet is limited [113Â115]. Acetylcholine and noradrenaline are locally released to act on cholinergic and adrenergic receptors. Acetylcholine stimulates insulin by enhancing Ca2+ release from intracellular depots, while activation of 2 -adrenergic receptors suppresses glucose-mediated insulin release [116] via hyperpolarization of the -cell membrane thus contributing to rapid adaptation of insulin secretion under conditions of hypoglycemia. On the contrary a stimulation of insulin secretion is elicited by activation of 2 -adrenergic receptors. To make the picture even more complex is the observation that a number of neurotransmitters. Also in this case, however, much of the information has been obtained with in vitro studies using murine islet models, while a careful profile of the human islet is still far from available. Moreover, the evaluation of the physiologic role of innervation of the pancreatic human islet poses a number of interpretation issues as stimulation or inhibition of the parasympathetic and sympathetic branches of the autonomic nervous system elicits an array of responses (metabolic changes, regional blood flow, release of multiple neurotransmitters and neuropeptides) that can ultimately result in a direct effect on the cell. By using more selective pharmacologic agents such as tyramine, a negative effect of sympathetic activation on insulin secretion induced by i. The islet innervation may play a main role in humans by mediating the cephalic phase of insulin secretion, that is, the rapid increase in insulin secretion occurring in the first couple of minutes upon initiation of the stimulus [120].
Purchase 75 mg sildenafil amex. What makes something "Kafkaesque"? - Noah Tavlin.
In type 2 diabetic patients with overt fasting hyperglycemia erectile dysfunction raleigh nc 25 mg sildenafil buy, a number of postbinding defects have been demonstrated erectile dysfunction treatment san antonio order 100 mg sildenafil with mastercard, including reduced insulin receptor tyrosine kinase activity erectile dysfunction 16 years old sildenafil 50 mg, insulin signal transduction abnormalities erectile dysfunction queensland buy sildenafil overnight, decreased glucose transport erectile dysfunction treatment massage proven 100 mg sildenafil, diminished glucose phosphorylation, and impaired glycogen synthase activity. When adipocyte storage capacity is exceeded, lipid "overflows" into muscle, liver, and cells, causing muscle/hepatic insulin resistance and impaired insulin secretion (reviewed in [311] and [312]). Lipid can also overflow into arterial vascular smooth cells, leading to the acceleration of atherosclerosis. Using 14 C-palmitate in combination with the insulin clamp technique [39], the antilipolytic effect of insulin has been shown to be markedly impaired in lean type 2 diabetic subjects, as well as in obese nondiabetic subjects [208]. After fatty acids enter the cell, they can be converted to triglycerides, which are inert, or to toxic lipid metabolites such as fatty acyl CoAs, diacylglycerol, and ceramide [4]. Using magnetic resonance spectroscopy, the intramyocellular triglyceride content has been shown to be increased in type 2 diabetic subjects [338,340]. Fatty acyl CoAs, which are known to inhibit insulin signaling [341,342], are also significantly increased in muscle in diabetic subjects [338,343]. However, there also is evidence that the mitochondrial defect is acquired, at least in part [349Â351]. Treatment of diabetic patients with pioglitazone markedly improves insulin sensitivity in association with a reduction in intramyocellular lipid and fatty acyl CoA concentrations. The decrement in muscle fatty acyl CoA content is closely related to the improvement in insulin-stimulated muscle glucose disposal [340,343]. Reduced intramyocellular fatty acyl CoA content with acipimox, a potent inhibitor of lipolysis, caused a similar improvement in insulin-mediated glucose disposal [338,339]. Increased intramyocellular levels of diacylglycerol [330,352] and ceramides [353,354] have been demonstrated in type 2 diabetic and obese nondiabetic subjects and shown to be related to the insulin resistance and impaired insulin signaling in muscle. Alpha cell and glucagon It long has been known that the basal plasma glucagon concentration is elevated in type 2 diabetic individuals [184Â186, 357,358]. There also is evidence that the liver is hypersensitive to the stimulatory effect of glucagon in hepatic gluconeogenesis [185]. The kidney: increased glucose reabsorption the kidney filters 162 g ([glomerular filtration rate = 180 L day-1] Ч [fasting plasma glucose = 900 mg L-1]) of glucose every day. Thus, an adaptive response by the kidney to conserve glucose, which is essential to meet the energy demands of the body, especially the brain and other neural tissues which have an obligate need for glucose, becomes maladaptive in the diabetic patient. Instead of dumping glucose in the urine to correct the hyperglycemia, the kidney chooses to hold on to the glucose. Even worse, the ability of the diabetic kidney to reabsorb glucose is augmented by an absolute increase in the renal reabsorptive capacity for glucose. The current epidemic of diabetes is being driven by the epidemic of obesity [369]. Porte and colleagues [370Â373] were amongst the first to demonstrate that, in rodents, insulin was a powerful appetite suppressant. Obese individuals, both diabetic and nondiabetic, have moderate-to-severe insulin resistance with compensatory hyperinsulinemia. Nonetheless, food intake is increased in obese subjects despite the presence of hyperinsulinemia which should suppress the appetite. Therefore, one could postulate that the insulin resistance in peripheral tissues also extends to the brain. After glucose ingestion, two hypothalamic areas with consistent inhibition have been noted: the lower posterior hypothalamus, which contains the ventromedial nuclei, and the upper posterior hypothalamus, which contains the paraventricular nuclei. In both of these hypothalamic areas, which are key centers for appetite regulation, the magnitude of the inhibitory response following glucose ingestion was reduced in obese, insulin-resistant, normal glucose-tolerant subjects, and there was a delay in the time taken to reach the maximum inhibitory response, even though the plasma insulin response was markedly increased in the obese group. Nonetheless, these results suggest that the brain, like other organs (liver, muscle, and fat) in the body, are resistant to insulin. Westermark P, Wilander E: the influence of amyloid deposits on the islet volume in maturity onset diabetes mellitus. Vaxillaire M, Froguel P: Monogenic diabetes in the young, pharmacogenetics and relevance to multifactorial forms of type 2 diabetes. Role of alterations in systemic, hepatic, and muscle lactate and alanine metabolism. Evidence for reduced insulin-dependent muscle glucose transport or phosphorylation activity in non-insulin-dependent diabetes mellitus. Insulin action and binding in isolated hepatocytes, insulin receptor structure, and kinase activity. A lesson in metabolic regulation inspired by the glucokinase glucose sensor paradigm. Identification of a protein kinase cascade of major importance in insulin signal transduction. Philosophical Transactions of the Royal Society London Series B: Biological Sciences 1999;354:485Â495. Kelley D, Mokan M, Mandarino L: Intracellular defects in glucose metabolism in obese patients with noninsulin-dependent diabetes mellitus. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Pathogenesis of type 2 diabetes mellitus 399 338 Bajaj M, Suraamornkul S, Romanelli A, et al. The risk variants are common in the population, have only a small effect on disease risk and together only explain a fraction of the heritability of the disease. The diabetic spectrum Diabetes is a group of metabolic diseases characterized by hyperglycemia resulting from defects in insulin secretion, insulin action, or both. The chronic hyperglycemia of diabetes is associated with long-term damage, dysfunction, and failure of different organs, especially the eyes, kidneys, nerves, heart, and blood vessels. These disease types could be divided further into subtypes possibly representing different disease mechanisms. The diabetes epidemic Worldwide prevalence figures estimate that there were 371 million diabetic patients in 2012 and more than 500 million are expected in 2030. In a situation of affluence most people tend to overconsume International Textbook of Diabetes Mellitus, Fourth Edition. One possible reason for this is that genetic selection has favored energy-preserving genotypes (so called thrifty genotypes); individuals living in an environment with unstable food supply could maximize their probability of survival by efficiently storing energy in times of surplus [2]. However, the exact proportion of the two, that is, the heritability, is difficult to determine and heritability estimates therefore vary in the range of 25Â80% between studies. Twin-studies have estimated the genetic component by comparing disease concordance in dizygotic twins with concordance in monozygotic twins. In these studies probandwise concordance rates (number of affected twins having a diabetic co-twin) for monozygotic twins vary between 34 and 100% [3Â6]. However, this statistic also varies depending on the cohort and population studied. There are also large differences between ethnic groups that seem to depend on genetic factors. While part of the observed ethnic variability could be attributed to environmental and cultural factors some of the variation seems to depend on genetic differences. One limitation of both twin and family studies is that they have a risk of overestimating heritability due to sampling and ascertainment errors. Concordant twins are more likely to participate in a study and the proportion of individuals with undiagnosed diabetes may differ compared to the general population. Another limitation is that family studies often cannot distinguish between the effect of genetic inheritance and the effects of a shared environment and the geneÂenvironment correlation that comes from nongenetic inheritance. There are many possible explanations for this including a role for the intrauterine environment in programming events later in life. Originally, disease-causing loci were identified primarily by linkage analysis, utilizing the long stretches of linkage in affected families. For loci identified by whole-genome association only loci reaching genome-wide significance (p < 5 Ч 10-8) are included. Finding that affected family members share a certain marker that is identical by descent. Linkage analysis has, however, been less useful for identifying genes causing complex diseases. This model has been the basis of a revolution in complex genetics by stimulating the development of tools for genetic association studies. The advantages of association lie in the larger number of individuals that can be collected for each study as well as the much higher resolution of the mapping. One disadvantage is the huge number of markers needed to perform mapping on a genome-wide level and association studies were therefore originally performed on small regions known to harbor candidate genes. The transcript expressed in adipose tissue has an extra exon B and a substitution of a proline for alanine at position 12 of this protein, which is seen in about 15% of the European population. It is well known that epinephrine excess can suppress insulin secretion and cause diabetes. Three of the studies shared results prior to publication and only considered positive results that were replicated in all studies. These very large studies have identified several new loci and confirmed the effect of many previously identified. Interestingly it seems like most associations found in one ethnic group also show some evidence of association in populations with other ethnicities. Therefore, additional fine mapping of the loci in even larger sample sets is often necessary. To do this cost-efficiently a Cardio-Metabochip has been developed for metabolic/cardiovascular gene mapping. Next-generation sequencing will provide even denser coverage of genetic variation. The GoT2D consortium aims to map lower frequency variants via low-coverage whole-genome sequencing, deep exome sequencing, and next-generation 2. Functions of associated genes Most identified diabetes loci have not been mechanistically tied to the disease. While loci are commonly referred to by the the genetics of type 2 diabetes 405 Table 26. Melatonin works as a chronobiotic factor, adjusting the timing of the biologic clock. Its receptors are present in the pancreas and melatonin is proposed to contribute to the nocturnal lowering of insulin in humans. The C allele of rs2943641 has been shown to be associated with insulin resistance and increased risk of diabetes. The risk allele is associated with decreased insulinogenic index and lower disposition index, suggesting a reduced capacity for insulin secretion in relation to insulin sensitivity. Since it was identified as a diabetes gene it has been shown to be important for several vital functions in the pancreatic islet, including pancreas development, determination of -cell mass, and maintenance of the secretory function of mature -cells. There are many possible explanations for the missing heritability, including assumptions made about the genetic architecture of the disease and the definitions of heritability. The estimations of heritability explained assumes that only additive affects determine disease risk and that the risk follows the liability threshold model, that is, the genetic and environmental effects sum up to form a normal distribution of liability and that disease arises in individuals surpassing a certain threshold in the distribution [58]. If these assumptions are not true, the estimate of heritability explained will not be correct. However, there are also many other potential explanations for the missing heritability: yet unmapped common variants, distorted parent-of-origin transmission of risk alleles, rare variants, structural polymorphisms. Some argue that disease risk is determined by a large number of very small additive effects from relatively common variants and that disease represents the extremes of a normal distribution [59] while others argue that complex diseases are rather collections of phenocopies caused by rare, often recent, mutations [60]. However, one should keep in mind that heritability can only be estimated from the most recent generations for which information on affection status is available, whereas most of the variants studied thus far are ancestral variants hundreds of generations old. We do not know whether these ancestral variants (which have modest effects and have escaped purifying selection) can really explain the diabetes epidemic we see in the most recent generations or whether this can be ascribed to rare variants with stronger effects. The rapid development of next generation sequencing tools has markedly facilitated discovery of rare variants. From being both extremely expensive and effort consuming, sequencing of the whole genome with appropriate coverage (approx. Whole-exome sequencing can be performed for less than $400 per sample and these figures are likely to decrease over the coming years. Applying these methods in families and large population studies will hopefully answer the questions about the role of rare variants in complex diseases. GeneÂgene and geneÂenvironment interactions GeneÂgene interactions, or epistasis, have been suggested as a possible explanation for difficulties in replicating genetic association in complex diseases [68]. The standard statistical methods used in association studies are usually limited to analysis of single marker effects and thereby do not account for interactions between markers. Previous attempts to study epistasis in complex diseases have focused on interactions between candidate regions [69,70]. The main problem when studying epistasis is power, since interaction between loci with modest effects is difficult to detect without extremely large sample sizes. Thorough studies in diabetes addressing epistasis using this approach are missing. Further, a recent paper by Eric Lander and coworkers provided compelling evidence that geneÂgene interaction can also contribute to missing heritability by causing "phantom heritability" that inflates the estimated narrow sense heritability of the trait [74]. Genetic variants could affect specific metabolic processes to make an individual more susceptible to the harmful effects of a poor diet but also personality traits that make an individual more or less likely to overconsume and live a sedentary lifestyle. It will however be a formidable task to identify the environmental triggers for most of the genetic variants increasing susceptibility to diabetes as the genetics of type 2 diabetes 409 this will require very large studies with precise information on diet, exercise, energy expenditure, and so on. Epigenetics the environment can also influence the expression of the genome, and ultimately the phenotype, via the epigenome. These modifications have the potential to be stable and heritable across cell divisions [75,76]. Changes to the phenotype can be at the level of the cell, tissue, or whole organism. Epigenetic mechanisms may, however, play a role in progression of the disease by inducing glucotoxicity in islets and predispose to diabetic complications.
Key references 988 Joint National Committee on detection erectile dysfunction doctor memphis cheap sildenafil 50 mg, evaluation and treatment of high blood pressure (988) erectile dysfunction electric pump cheap sildenafil 75 mg fast delivery. The 988 report of the Joint National Committee on detection erectile dysfunction age 40 proven sildenafil 50 mg, evaluation and treatment of high blood pressure erectile dysfunction doctors in arizona purchase sildenafil 100 mg fast delivery. Means at each examination and inter-examination consistency of specified characteristics: Framingham Heart Study: 30-year follow-up erectile dysfunction qarshi generic 75 mg sildenafil overnight delivery. Prognostic implications of baseline electrocardiographic features and their serial changes in subjects with left ventricular hypertrophy. Reduction of cardiovascular risk by regression of electrocardiographic markers of left ventricular hypertrophy by the angiotensin-converting enzyme inhibitor ramipril. Blood cholesterol and vascular mortality by age, sex, and blood pressure: a meta-analysis of individual data from 6 prospective studies with 55,000 vascular deaths. Sundstrom J, Arima H, Jackson R, for the Blood pressure Lowering Treatment Trialists Collaboration (204). Effects of blood pressure reduction in mild hypertension: A systematic review and meta analysis. As early as the 970s, the significance of hypertension in the pathogenesis of atherothrombotic stroke was established, and newer studies continue to confirm the benefit achieved from blood pressure reduction in the prevention of cardiovascular and cerebrovascular diseases. For example, hypertension is associated with abnormalities in coagulation, platelets, and the endothelium, leading to a prothrombotic or hypercoagulable state. The latter may explain why despite the blood vessels being exposed to high pressures, the main complications of hypertension-that is, heart attacks and strokes-are thrombotic rather than haemorrhagic. The changes in the retina (hypertensive retinopathy) are the most widespread early changes that are seen and that have been described. However, it is now thought that these classifications do not correlate well with the severity of hypertension and progression, and a new simpler two-grade classification of non-malignant versus malignant retinopathy has been proposed. The most common ocular manifestation of hypertension is narrowing of the retinal arterioles. In young patients, the arterioles may constrict due to autoregulation as described in the following text. In older patients, luminal fibrosis and vessel rigidity prevent the same degree of narrowing. At points where the retinal arteriole crosses over the retinal venule, compression of the vein may cause the appearance of arteriovenous nicking. Under normal circumstances, there are many feedback mechanisms that maintain retinal flow despite changes in blood pressure. Retinal vessels have the ability to maintain a constant blood flow despite changes in perfusion pressures by either vasodilation or vasoconstriction. However, with hypertension, there is a breakdown of this mechanism, due to changes in endothelial-derived molecules (endothelins, thromboxane A2, etc. This breakdown of the autoregulation leads to other changes such as oedema and fibrosis. In malignant hypertension, the changes seen include papilloedema as well as hard and soft exudates which are due to severe vasospasm of the vessels in response to the high pressures (as seen in malignant hypertension), leading to necrosis and focal leakage from the precapillary arterioles that lie deep in the retina. Fundoscopy showing malignant hypertensive changes with papilloedema, haemorrhages, infarcts (cotton wool spots), and arteriolar constriction 8. The Framingham Heart Study was one of the first to demonstrate that attention and memory measures are inversely related to blood pressure levels and duration of hypertension. Hypertension is a risk factor for atherosclerosis, stroke, or cerebral infarction, which in turn may cause cognitive decline. In the absence of an overt cerebrovascular accident or stroke, cognitive impairment may be a result of occlusion of microvasculature. Patients with hypertension have smaller volumes of thalamic nuclei and larger volumes of cerebrospinal fluid in the cerebellum and temporal lobes. The Systolic hypertension in Europe Study (Syst Eur) was one of the first studies to demonstrate a protective effect of antihypertensive therapy on the development of cognitive impairment. Antihypertensive therapy has also been shown to reduce the incidence of white matter changes on magnetic resonance imaging. It is epidemiologically more common to see renal failure leading to hypertension, but the converse is controversial, except in malignant hypertension, where progressive deterioration of renal function has been demonstrated. Here as well, the glomerular vessels autoregulate the blood flow by vasoconstriction or vasodilatation depending on the perfusion pressures, to keep the actual perfusion at the glomerulus constant. Prolonged high perfusion pressures can lead to significant vasoconstriction, which can then cause localized damage to the glomeruli. This can cause necrosis of the glomeruli leading to microalbuminuria, which could lead to significant proteinuria if the disease is not treated. Renal failure in the absence of the malignant phase could also be an effect of atherosclerosis affecting the renal arteries, leading to underperfusion. The reversal of microalbuminuria with the strict treatment of hypertension has been shown to improve cardiovascular events. There are many factors involved and no factor, on its own, can be singled out as the most important one. It is an interaction of these various factors that causes the effects noted in the various organ systems. Changes in the endothelium the changes in the endothelium in hypertension are widespread and well established. This activated endothelium causes increased production of procoagulant and inflammatory agents and is associated with a shift of the actions of the endothelium towards reduced vasodilation, a proinflammatory state, and prothrombic properties. With constant exposure to this high-pressure flow, and as a result of the inflammatory mediators and the effects of the neutrophils and platelets that have got adhered to the endothelium, it then starts to become dysfunctional. Endothelial dysfunction is associated with different cardiovascular risk factors such as ageing, postmenopausal status, hypercholesterolemia, diabetes mellitus, smoking, hyperhomocysteinaemia, and hypertension. Combination of these risk factors has been shown to lead to further deterioration of the endothelial-dependent vasodilatation. Therefore, endothelial dysfunction is now considered a relevant mechanism that promotes atherosclerosis and thrombosis. Conceivably, endothelial function can also be used as an intermediary end point in the treatment of hypertension. Although the condition is associated with high pressures, it is commonly complicated by thrombotic events such as ischaemic strokes and myocardial infarctions. Platelet activation fulfills the third criterion of alteration to the blood constituents. Many changes in the coagulation and fibrinolytic pathways of hypertensive patients have been documented. Fibrin D-dimer, an index of fibrin turnover and 53 thrombogenesis, has also been shown to be raised in hypertensives. Plasma fibrin D-dimer levels are predictive of both arterial thrombotic events and postoperative thrombosis. In hypertension, it has been shown that there is a decrease in the levels of the collagenases and an increase in the levels of their inhibitors, leading to an increase in the laying down of the extracellular matrix. This is especially important, as it has been shown that high plasma renin activity in hypertensive patients is associated with increased risk for coronary events and stroke. The microcirculation is important in regulation of peripheral vascular tone and it is altered in hypertension. It should be noted that other comorbidities, such as atrial fibrillation, diabetes, and congestive heart failure are more common in hypertensive patients, and are known to activate platelets. Conceivably, some consider that the atherosclerotic lesions that are seen with hypertensive patients could act as a nidus for further platelet activation. However, it has been suggested that vessel wall thickening and a response of the muscular wall of the vessels to increased flow and blood pressures causes this rarefaction. This vasoconstriction of the peripheral vessels causes increased peripheral resistance, which then perpetuates the increased blood pressure and sets up a vicious cycle. Pulse pressure, mean arterial pressure, and pressure wave, along with cardiac output, also affect microcirculation. It has been suggested that increase in angiogenic factors is a compensatory mechanism as a result of tissue hypoxia due to the rarefaction of the microcirculation. The increase in thrombogenesis, impaired fibrinolysis, and platelet activation could be responsible for macrovascular diseases such as strokes and peripheral vascular disease, whilst all the changes together could result in increased atherogenesis. The main aim in the management of a patient with hypertension is the prevention of cardiovascular and cerebrovascular complications. The importance of good blood pressure control towards this end cannot be overemphasized. Medical literature is full of studies demonstrating the clinical benefits of good blood pressure control in preventing cardiovascular and cerebrovascular endpoints, irrespective of the agent used. Whilst it has been shown convincingly that the use of antihypertensive agents reduces cardiovascular and cerebrovascular complications and that they reverse endothelial and platelet activation in hypertension, a direct correlation between the improvement in endothelial and platelet activation and a decrease in cardiovascular endpoints has not been shown. More studies are needed to fully understand the different mechanisms involved in the pathogenesis of target organ damage in hypertension and in devising strategies to prevent them. White coate hypertension as defined by ambulatory blood pressure monitoring and subclinical cardiac organ damage: a meta-analysis. The endothelium as prognostic factor and therapeutic target: what criteria should we apply? Muiesan M, Salvetti M, Rizzoni D (203) Resistant hypertension and target organ damage. The prothrombotic state in hypertension and the effects of antihypertensive treatment. Target organ damage in hypertension: pathophysiology and implications for drug therapy. Left atrial involvement: 3 points (terminal negativity of P wave in V mm with duration 0. Although in vitro studies appear to show a correlation between the two, in vivo studies have not been that definite. Insulin has trophic effects, and insulin resistance is more common among hypertensive patients than in normotensive subjects. Insulin resistance is associated with an attenuated therapeutic response to antihypertensive drugs. In newly diagnosed hypertensives, fasting plasma leptin levels determine myocardial wall thickness independently of 24-hour ambulatory blood pressure levels. This relationship is independent of other factors such as age, gender, blood pressure, smoking, cholesterol, and diabetes. Concentric hypertrophy appears to carry the highest risk and eccentric hypertrophy an intermediate risk. The reason for this difference in prognosis for the different geometric patterns is not certain. This pattern is associated with severe haemodynamic and structural abnormalities and would therefore represent a more unfavorable pattern. Almost six decades ago it was first demonstrated that cardiac hypertrophy could be reversed by nephrectomy in spontaneously hypertensive rats. After adjusting for various variables, there was a significant difference among the different medication classes. Endothelin is known to be a potent vasoconstrictor that is implicated in the pathogenesis of hypertension. An aggressive therapeutic management directed at achieving a blood pressure reduction over a 24-hour period and a strict control of concomitant modifiable risk factors seem mandatory in these high-risk patients. Prevalence of left-ventricular hypertrophy in hypertension: an updated review of echocardiographic studies. A meta-analysis of the effects of treatment on left ventricular mass in essential hypertension. Echocardiographic criteria for left ventricular hypertrophy: the Framingham Heart Study. Relationship between left ventricular systolic function to persistence or development of electrocardiographic left ventricular hypertrophy in hypertensive patients: implications for the development of new heart failure. Accuracy of electrocardiography in diagnosis of left ventricular hypertrophy in arterial hypertension: systematic review. Therefore it comes as no surprise that hypertension is the most prevalent concomitant condition in patients with atrial fibrillation in Europe and North America and they are thought to be inextricably linked. Therefore the importance of reversing or prevention of new atrial fibrillation should not be underestimated. These include structural changes, neuro-endocrine activation and myocardial fibrosis. Changes in calcium handling due to micro re-entrant circuits result in reduction of effective atria contraction leading to stasis of blood within atria and therefore promoting haemostasis with subsequent increased risk of thrombo-embolic stroke. The fibroblasts of the enlarged atria subsequently differentiate into myofibroblasts that enhance connective tissue deposition and fibrosis. These include angiotensin-converting enzyme inhibitors and angiotensin receptor blockers, beta blockers, and calcium channel blockers. There is at present a lack of data suggesting the prevention of new-onset atrial fibrillation. The co-existence of hypertension and atrial 69 fibrillation has been reported to double the risk for all of the forementioned. This emphasizes the importance of optimizing medical treatment strategies of hypertensive patients, thereby minimizing potential complications. Beta blockers are effective for rate control with certain beta blockers maintaining sinus rhythm. Clinical study to investigate the predictive parameters for the onset of atrial fibrillation in patients with essential hypertension. The Working Group on Hypertension, Arrhythmias and Thrombosis of the European Society of Hypertension (202).
References
- Leacche M, Unic D, Goldhaber SZ, et al: Modern surgical treatment of massive pulmonary embolism: Results in 47 consecutive patients after rapid diagnosis and aggressive surgical approach, J Thorac Cardiovasc Surg 129:1018, 2005.
- Efremidis M, Pappas L, Sideris A, et al. Management of atrial fibrillation in patients with heart failure. J Card Fail 2008;14:232.
- Debaene B, Plaud B, Dilly MP, et al: Residual paralysis in the PACU after a single intubating dose of nondepolarizing muscle relaxant with an intermediate duration of action, Anesthesiology 98(5):1042-1048, 2003.
- Olson L, Eckerdal O, Hallonsten AL, et al. Craniomandibular function in juvenile chronic arthritis. A clinical and radiographic study. Swed Dent J 1991;15:71-83.
- Bolling SF, Su TP, Childs KF, et al: The use of hibernation induction triggers for cardiac transplant preservation, Transplantation 63(2):326-329, 1997.
- Tritschler HJ, Andreetta F, Moraes CT, et al. Mitochondrial myopathy of childhood associated with depletion of mitochondrial DNA. Neurology 1992;42:209.