

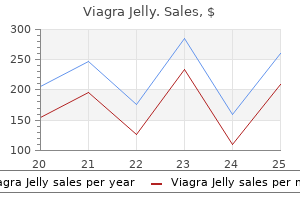
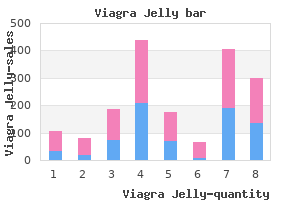
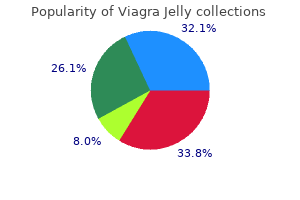
Viagra Jelly
| Contato
Página Inicial
Anders A.F. Sima, M.D., Ph.D.
- Professor of Pathology and Neurology
- Wayne State University School of Medicine
- Detroit, MI
In primary active transport impotence gandhi proven 100 mg viagra jelly, a transporter protein is directly involved with the transport of solutes across the membrane erectile dysfunction shot treatment buy 100 mg viagra jelly otc. Second erectile dysfunction medication injection buy 100 mg viagra jelly mastercard, the protein binds sodium from the intracellular compartment and potassium from the extracellular compartment erectile dysfunction hypertension medications order viagra jelly without a prescription. This change in structure allows three sodium ions to be transferred from the intracellular fluid to the extracellular fluid while at the same time moving two potassium ions from the outside of the cell to the inside of the cell impotence def purchase 100 mg viagra jelly with amex. In this way the cell maintains concentration gradients of both sodium and potassium, thereby preventing the cell from reaching chemical equilibrium. Similar active transport systems move other combinations of ions across the plasma membrane. Secondary active transport involves ion pumps, but the activity of these pumps serves to facilitate movement of solutes other than the ions. To understand this process it is important to note that glucose is transported into the cell on its apical surface (facing the lumen of the intestine) and then out of the cell on its basal surface (facing interstitial space). The symporter moves Na and glucose into the cell down the concentration gradients of these solutes. Still, the movement of such molecules into and out of the cell is necessary to maintain homeostasis. Macromolecules such as proteins are transported across the plasma membrane in small, bubble-like structures called vesicles. Vesicles are surrounded by membranes and can be transported throughout the cytosol as well as across the cell membrane. Endocytosis is the process by which cells engulf and take in substances from the extracellular compartment. Pinocytosis ("cell drinking") is a form of endocytosis involving the ingestion of fluids. Both phagocytosis and pinocytosis are nonspecific and constitutive (always taking place). A third type of endocytosis is triggered by specific chemicals and is called receptor-mediated endocytosis. Exocytosis refers to the cellular release of substances via vesicular transport; namely, secretion. In regulated (non-consitutive) exocytosis, also known as secretion, the entire process is triggered by certain chemicals called secretagogues. The calcium then interacts with a specific calcium-sensing protein that promotes docking and fusion of the vesicle with the plasma membrane. This may involve changing size or shape and/or movement of cell appendages such as flagella or cilia. At this point it is only necessary to emphasize the fact that movements can be vital to homeostasis and that such movements require energy. All cell movements involve so-called motor proteins that make up part of the cytoskeleton. Movement is typically the result of complex chemical interactions that cause one protein to interact with and move another protein; for example, shortening of muscle cells (myofibers) is the result of complex chemical interactions that cause actin molecules to slide along myosin molecules. As noted in the previous section, some of these are also involved with cell movement. Many cells produce other types of functional proteins that reside on the exterior surface of the plasma membrane. The aforementioned examples support the idea that the manufacturing of proteins is vital to overall regulation of homeostasis. Protein synthesis is a major source of energy consumption in cells, especially during growth of cells and tissues. Before we delve into the details of protein synthesis it is necessary bring up another theory that is fundamental to modern biology: gene theory. A detailed understanding of the molecular aspects of these processes is not necessary for this introductory consideration of human anatomy and physiology. We will therefore review only the major steps in order to shed light on subsequent discussions of how these processes regulate homeostasis. The two strands, running in opposite directions, are joined by pairs of the bases. The structure is analogous to that of a ladder; that is, the backbones form the side rails and the joined base pairs form the rungs. The entire molecule has a right-handed twist to give it the structure of a "double helix. The exposed sequence of these purines and pyrimidines make up the so-called genetic code-a message that provides information for synthesizing a particular protein. For our purpose it is only necessary to consider the major steps in transcription. Proteins are made up of various combinations of 22 amino acids, each of which corresponds to at least one codon. As the protein emerges from the ribosome the ribosome docks with the membrane of the endoplasmic reticulum, and the chain of amino acids penetrates and elongates into the matrix of the endoplasmic reticulum. At this point the emerging protein is clipped away from the ribosome and remains in the matrix of the endoplasmic reticulum. The proteins that enter the endoplasmic reticulum are chemically modified and are eventually transported to the Golgi complex via vesicular transport. Cell function is regulated by two main mechanisms: 1) gene regulation and 2) enzyme regulation. Cells of different tissues express different genes that allow them to perform their specific functions. Whether or not a cell produces a particular protein depends on whether or not the gene that codes for the protein is expressed. Differences in gene expression among different cell types have been attributed to regulation of the gene promoters-mechanisms that activate or repress transcription of genes. Enzyme regulation refers to changes in activity of certain proteins that regulate rates of chemical reactions in cells. Transcription and translation are necessary to produce such proteins, but the activity of enzymes is often regulated by certain chemical substances. Enzyme inhibitors bind to the enzyme and prevent it from acting, whereas enzyme activators interact with the protein to enhance its activity. It is important to bear in mind that changes in cell activity can be attributed to gene regulation or enzyme regulation, or a combination of the two processes. Also note that gene expression is a prerequisite for enzyme regulation; that is, the enzyme must be synthesized before it can be regulated. Liver cells, for example, have one of the shorter turnover rates; that is, 300400 days. The important point is that the continual reproduction and growth of cells is required to sustain functions of most tissues. Cell reproduction refers to the replication of a cell, whereas cell growth involves a change in the size of a cell. In order to maintain homeostasis of the skin, the body must continually produce new skin cells. We will study this process in detail in Chapter 4, but we can refer to it now to differentiate between cell reproduction, growth, and differentiation. The deepest of these consists of stem cells; that is, cells that have the potential to become any type of cell. The stem cells of skin are small, cuboid shaped, and have a high rate of mitosis (cell division); that is, the formation of two identical offspring cells from a so-called parent cell. The high rate of mitosis in this layer results in an increase in cell numbers, a process commonly referred to as hyperplasia. The change in size is a form of cell growth called hypertrophy, while the change in structure and function is referred to as differentiation. The growth and differentiation of skin cells occur as the cells move upward, thereby displacing the oldest cells that are continually shed. The changes caused by differentiation eventually cease, and the cell undergoes a process called apoptosis, or programmed cell death. These dead cells make up several layers of skin cells and provide a barrier that protects the internal environment. The balance between cell reproduction and cell death sustains the skin in a homeostatic state. The cell cycle is a repeating sequence of events that are necessary for a cell to reproduce. Continuous growth of the cell characterizes the interphase, whereas mitosis refers to the duplication of the cell into two identical offspring cells. The process consists of four phases that refer to the shapes and orientations of chromosomes. Meiosis is a second form of cell reproduction that is unique to sexually reproducing organ- isms. The process has also been called reduction division because meiosis involves cell division and reduction of the chromosome number. Gametes have only half the number of chromosomes as the other cells of the body. Describe all of the intracellular mechanisms whereby hormone A might increase concentrations of albumin. As with macroscopic anatomy, the goal of histology is to associate structure with function. Unlike gross anatomy, histology requires specialized equipment to make these associations. Various types of microscopes are required to visualize the microanatomy of cells and tissues. The light microscope is the primary tool for learning the microanatomy of tissues. This article provides photomicrographs (photographs of microscopic images) of various tissue types as viewed with a light microscope. It is also necessary to stain the tissue so cells and noncellular substances can be clearly visualized. Samples range in size but are typically several millimeters thick and no larger than 1 cm in length and width. Fixation preserves the structure of the tissue by: 1) terminating cell metabolism; 2) preventing enzymatic degradation (autolysis) of cells and tissues; 3) killing pathogenic microorganisms, and 4) altering structural proteins to harden the tissue. Once the tissue sample is fixed it can be embedded in a solid block of paraffin to facilitate sectioning. The specimen is then placed into a small form and melted paraffin is poured over it. After cooling, a hardened block of paraffin is trimmed to an appropriate size and mounted on a microtome, a machine that is designed to make precise, ultra-thin slices with a razor-sharp knife. Each slice is mounted onto a glass slide and then subjected to the staining process. It is therefore necessary to stain the tissue in order to highlight its microanatomy. Before staining is possible the paraffin is dissolved away from the sample and the tissue is rehydrated. A common staining method involves two stains: hematoxylin, which highlights only some structures, and eosin, a counterstain that highlights other structures. The combination of the two stains produces a section in which various structures are easily distinguishable. After completion of the staining process a cover slip is placed over the specimen. One of the major frustrations stems from the fact that light microscopy involves two-dimensional images of three-dimensional objects. This means that when viewing a slice of tissue, the viewer sees only two dimensions and must imagine the third one. To develop this type of spatial ability it is necessary to examine various sections of the tissue; for example, median, horizontal, and frontal sections. It is also important to keep in mind that spherical or tubular structures can appear in many different shapes depending on how they are sectioned. Artifacts can arise from errors in collecting, mounting, sectioning, and staining. The ability to distinguish between a cell nucleus and a dark spot of stain takes time to develop and requires experience and familiarity with the preparation process. A tissue is a highly organized aggregation of cells that interact in a cooperative manner to perform certain functions. The organization of cells therefore provides insight into how they cooperate to perform the functions of organs. There are only four basic tissue types: · Epithelial · Connective · Muscle · Nervous Each of these tissue types conforms to a set of structural and functional. Within each category there are subtypes that are characterized by particular cell types and extracellular materials. Identification of tissue types requires more practice and experience than you will acquire in an introduction to anatomy. A reasonable goal is to learn the major structural features of the different cell arrangements that define the four major classes of tissues. Epithelial cells are always in close proximity to each other and are typically joined by specialized membrane proteins called junctions. This, of course, includes the skin, but it also includes the linings of the body cavities. Each of these surfaces is an external portion of the body or is continuous with the exterior. The outermost layer of epithelial cells form the free surface; that is, the most superficial layer. The deepest layer of epithelial cells rests on a thin layer of connective tissue called the basement membrane.
Mental retardation is common and can be attributed to many genetic and environmental causes other uses for erectile dysfunction drugs 100 mg viagra jelly mastercard. The newly formed brain consists of three parts: the prosencephalon impotence 2 100 mg viagra jelly buy fast delivery, the mesencephalon trimix erectile dysfunction treatment buy discount viagra jelly on-line, and the rhombencephalon causes of erectile dysfunction in 30s order viagra jelly 100 mg visa. The prosencephalon later subdivides into the secondary prosencephalon and the diencephalon discussing erectile dysfunction doctor order viagra jelly master card, and the rhombencephalon forms the metencephalon and myelencephalon. Among the glial cells, radial glial cells act as guidewires for the migration of neurons from their sites of origin to definite layers in the brain. Neuroblasts in the intermediate zone (future gray matter) send out processes that collect principally in the marginal zone (future white matter). The neural tube is also divided into a dorsal alar plate and a ventral basal plate. The basal plate represents the motor component of the spinal cord, and the alar plate is largely sensory. Further influences of shh, produced by the notochord and the floor plate, result in the induction of motoneurons in the basal plate. This structure is reflected in the rhombomeres and molecularly in the patterns of expression of homeobox-containing genes. Neurons and their processes developing within the rhombomeres follow specific rules of behavior with respect to rhombomere boundaries. Nerve processes growing from the spinal cord react to external cues provided by the environment of the somites. Neurons and neural crest cells can readily penetrate the anterior but not the posterior mesoderm of the somite. This growing tip continually samples its immediate environment for cues that guide the amount and direction of axonal growth. The motor component of a peripheral nerve is joined by the sensory part, which is based on neural crestderived cell bodies in dorsal root ganglia along the spinal cord. Axons and dendrites from the sensory cell bodies penetrate the spinal cord and grow peripherally with the motor axons. Connections between the nerve and end organs are often mediated through trophic factors. Typically, sympathetic neurons are adrenergic, and parasympathetic neurons are cholinergic. The normal choice of transmitter can be overridden, however, by environmental factors so that a sympathetic neuron can secrete acetylcholine. During the fetal period, growth in the length of the spinal · · · · · · cord lags behind that of the vertebral column, thus pulling the nerve roots and leaving the end of the spinal cord as a cauda equina. Within the brain, the myelencephalon retains an organizational similarity to the spinal cord with respect to the tracts passing through, but centers that control respiration and heart rate also form at the site. The metencephalon contains two parts: the pons (which functions principally as a conduit) and the cerebellum (which integrates and coordinates many motor movements and sensory reflexes). In the cerebellum, which forms from the rhombic lips, the gray matter forms on the outside. The ventral part of the mesencephalon is the region through which the major tracts of nerve processes that connect centers in the cerebral cortex with specific sites in the spinal cord pass. The dorsal part of the mesencephalon develops the superior and inferior colliculi, which are involved with the integration of visual and auditory signals. Many important nuclei and integrating centers develop in the diencephalon, among them the thalamus and pineal body. The secondary prosencephalon gives rise to the telencephalic vesicles and the hypothalamus, including the neurohypophysis and optic chiasms. Similar to the cerebellum, the telencephalon is organized with the gray matter in layers outside the white matter. Neuroblasts migrate through the white matter to these layers by using radial glial cells as their guides. The cranial nerves are organized on the same fundamental plan as the spinal nerves, but they have lost their regular segmental pattern and have become highly specialized. Many congenital malformations of the nervous system are based on incomplete closure of the neural tube or associated skeletal structures. In the spinal cord, the spectrum of defects ranges from a widely open neural tube (rachischisis) to relatively minor defects in the neural arch over the cord (spina bifida occulta). Neural function appears in concert with the structural maturation of various components of the nervous system. During successive weeks, the reflex movements become more complex, and spontaneous movements appear. Final functional maturation coincides with myelination of the tracts and is not completed until many years after birth. What molecule produced by the notochord is instrumental in inducing the floor plate of the neural tube An infant with a tuft of hair over the lumbar region of the vertebral column undergoes surgery for a congenital anomaly in that region. During surgery, it was found that the dura and arachnoid layers over the spinal cord were complete, but that the neural arches of several vertebrae were missing. Complete failure of the neural tube to close in the region of the spinal cord is a. Pregnant women typically first become aware of fetal movements during what month of pregnancy In the early days after birth, an infant does not pass fecal material and develops abdominal swelling. What is the likely appearance of the spinal cord and brachial nerves in an infant who was born with the congenital absence of one arm (amelia) The cell biology of neurogenesis: toward an understanding of the development and evolution of the neocortex. Development of coherent neuronal activity patterns in mammalian cortical networks: common principles and local heterogeneity. Noses and neurons: induction, morphogenesis, and neuronal differentiation in the peripheral olfactory pathway. A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube. Getting axons onto the right path: the role of transcription factors in axon guidance. Pattern formation in the vertebrate neural tube: a sonic hedgehog morphogen-regulated transcriptional network. Parasympathetic neurons originate from nerve-associated peripheral glial precursors. Siah regulation of Pard3A controls neuronal cell adhesion during germinal zone exit. Morphogen to mitogen: the multiple roles of hedgehog signaling in vertebrate neural development. Slit-mediated repulsion is a key regulator of motor axon pathfinding in the hindbrain. Mechanisms regulating the development of the corpus callosum and its agenesis in mouse and human. The initial appearance of the cranial nerves and related neuronal migration in staged human embryos. The timing and sequence of appearance of neuromeres and their derivatives in staged human embryos. Hoxa2- and rhombomere-dependent development of the mouse facial somatosensory map. Otx dose-dependent integrated control of anteroposterior and dorso-ventral patterning of midbrain. Oligodendrocyte precursors migrate along vasculature in the developing nervous system. Wnt won the war: antagonistic role of Wnt over shh controls dorso-ventral patterning of the vertebrate neural tube. Integrity of developing spinal motor columns is regulated by neural crest derivatives at motor exit points. How does Fgf signaling from the isthmic organizer induce midbrain and cerebellum development Sonic hedgehog functions through dynamic changes in temporal competence in the developing forebrain. Among the new transcription factors upregulated in specified neural crest precursor cells are snail-1,-2 (formerly called slug), Twist, and Foxd-3, which are instrumental in allowing the neural crest cells to undergo an epitheliomesenchymal transformation. These cells then break free from the neural epithelium and then migrate away as mesenchymal cells. Neural crest cells break free from the neural tube in the trunk at the level of the last-formed somite or the neural plate in the head by changing their shape and properties from those of typical neuroepithelial cells to those of mesenchymal cells. These molecules remain downregulated during migration, but after neural crest cells have completed their migrations and have differentiated into certain structures. In the head, where closure of the neural plate has not yet occurred, neural crest cells must penetrate the basal lamina underlying the neural plate. This is accomplished by the production of enzymes that degrade components of the basal lamina and by sending out processes that penetrate the basal lamina. In the trunk, neural crest cells do not leave the neuroepithelium until after the neural tube has formed. They do not, however, have to contend with penetrating a basal lamina because the dorsal part of the neural tube does not form a basal lamina until after emigration of the crest cells. The neural crest, the existence of which has been recognized for more than a century, forms an exceptionally wide range of cell types and structures, including several types of nerves and glia, connective tissue, bones, and pigment cells. Its importance and prominence are such that the neural crest has often been called the fourth germ layer of the body. Not until adequate methods of marking neural crest cells became available-first with isotopic labels and subsequently with stable biological markers, monoclonal antibodies, intracellular dyes, and genetic markers-did the neural crest become one of the most widely studied components of the vertebrate embryo. More recently, emphasis has shifted to studies on the mouse, especially for dissecting molecular controls, but it appears that most of the information on the biology of the neural crest derived from birds can be applied to mammalian embryos. Some important syndromes and malformations are based on abnormalities of the neural crest. Tracing the history of the neural crest in any region involves consideration of the following: (1) its origin, induction, and specification; (2) epithelial-tomesenchymal transformation and emigration from the neural tube; (3) migration; and (4) differentiation. Each of these phases in the development of the generic neural crest is covered before neural crest development in specific regions of the body is considered. In addition to neural crest precursors, the neural plate border contains several types of progenitor cells, such as progenitors of the ectodermal placodes in the anterior region. In response to these inductive signals, cells at the border of the neural plate activate genes coding for several transcription factors, including Msx-1,-2, Dlx-5, and Pax-3/Pax-7. They also activate another set of genes (Foxd-3, Sox-10, and Ets-1), which specify the neural crest progenitor cells within the neural plate border. In this environment, the cells undergo extensive migrations along several well-defined pathways. These migrations are determined by intrinsic properties of the neural crest cells and features of the external environment encountered by the migrating cells. One of its main functions is to prevent premature differentiation of the migrating cells. Neural crest migration is influenced by a variety of molecules residing in the extracellular matrix. Although the presence of a basal lamina can inhibit their emigration from the neural tube, neural crest cells often prefer to migrate along basal laminae, such as those of the surface ectoderm or neural tube, after they have left the neural tube. Components of the extracellular * Snail-2 is also expressed during gastrulation by cells of the epiblast after they have entered the walls of the primitive streak and are about to leave as mesenchymal cells of the mesoderm germ layer. Overall, intrinsic factors are heavily involved in organizing the migrating streams of cells. During their migratory phase, neural crest cells are exquisitely sensitive to guidance molecules, most of which are inhibitory. Among the most important of these guidance molecules are the ligand/receptor pairs Robo/Slit, Neuropilin/Semaphorin, and Ephrin/Eph (see Table 11. Migratory neural crest cells extend protrusions that both test the environment and are part of the propulsive mechanism. This mechanism acts as a brake when the cells encounter an inhibitory environment, but it is also involved in their forward propulsion. In a migrating stream of neural crest cells, contact with the cells behind also results in the pulling at the trailing edge of the cells, thus resulting in a net forward motion of the leading cells. Specific examples of the environmental control of neural crest cell migrations are given later in this chapter. Much remains to be learned about what causes neural crest cells to stop migrating, but often they stop migrating in areas where repulsive signals are low. What controls their differentiation is a principal question of neural crest biology. According to one, all neural crest cells are equal in developmental potential, and their ultimate differentiation is determined by the environment through which they migrate and into which they finally settle. The other hypothesis suggests that premigratory crest cells are already programmed for different developmental fates, and that certain stem cells are favored, whereas others are inhibited from further development during migration. More recent research indicates that the real answer can be found somewhere between these two positions. Increasing evidence suggests that among migrating neural crest cells is a mix of cells the fate of which has been predetermined within the neural tube and cells the ultimate phenotype of which depends on environmental influences. A correlation exists between the time of migration of neural crest cells from the neural tube and their developmental potential.
Buy cheap viagra jelly 100 mg. Erectile Dysfunction Mental Health & Wellbeing | Kati Morton | Kati Morton.
During development experimental erectile dysfunction drugs discount viagra jelly 100 mg otc, more proximal segments differentiate first erectile dysfunction after 80 viagra jelly 100 mg line, followed successively by the more distal segments erectile dysfunction talk your doctor buy generic viagra jelly 100 mg line. In the early limb bud erectile dysfunction drugs pictures buy viagra jelly 100 mg on line, the proximal mesenchymal cells are exposed to a high concentration of retinoic acid because they are near the source (somites) erectile dysfunction with diabetes 100 mg viagra jelly order mastercard, and they differentiate into tissues of the stylopod. While the limb bud grows out, the remaining undifferentiated cells are exposed to a lesser concentration of retinoic acid because outgrowth has taken them farther from the source of retinoic acid. Thus those remaining mesenchymal cells in later limb buds first differentiate into the zeugopodial segment and finally, in the late limb bud, into the autopodial segment. Using many of the same components as those involved in somitogenesis such as hairy and the delta/notch system, the mesenchymal cells in the developing limb may respond to oscillations of the clock mechanism in forming the skeletal components of the major proximodistal segments. Because of these exclusive expression regions, it was originally assumed that these two genes determined the identity of the forelimb and hindlimb. The main functions of Tbx-4 and Tbx-5 appear to be the initiation of development in a limb-specific manner. Pitx-1, which is also expressed in the hindlimb, may play a more important role than Tbx-4 as a determinant of hindlimb identity. When the limb bud takes shape, its further development depends to a great extent on the actions of three signaling centers, one for each of the cardinal axes of the limb. First, it antagonizes Gli-3, confining Gli-3 activity to the anterior part of the limb bud, where it represses the expression of posterior limb genes. Organization of the dorsoventral axis of the limb begins when the dorsal ectoderm produces the signaling molecule, Wnt-7a, which stimulates the underlying limb bud mesenchyme to express the transcription factor, Lmx-1b, a molecule that imparts a dorsal character to the mesoderm underlying the dorsal ectoderm. This sequence represents a second wave of Hox gene expression after that involved in the initiation of limb development. Shh stimulates the expression of the Hox genes in the limb, and Gli-3 is involved in confining Hox gene expression to the more posterior parts of the limb bud. Studies on mice and the analysis of certain human mutants have shown that certain defects in limb regions correspond to absent expression of specific Hox gene paralogs. An interesting but little explored question in the development of many structures is what causes development to cease. If the intervening wedge of formerly shhproducing cells is removed from the distal tip of the mature limb bud, the more anteriorly located mesodermal cells are again exposed to above-threshold concentrations of shh, and they can again produce gremlin. The result is the formation of digits with more than the normal number of phalangeal segments. Cell Death and Development of Digits Although it may seem paradoxical, genetically programmed cell death (apoptosis) is important in the development of many structures in the body. Experiments on avian embryos showed that, to a certain stage, mesodermal cells scheduled to die could be spared by transplanting them to areas in which cell death did not normally occur. After a certain time, however, a "death clock" was set (an example of determination), and the cells could no longer be rescued. Instead of remaining continuous around the entire apex of the limb, the ridge begins to break up, leaving intact segments of thickened ridge epithelium covering the emerging digital rays (cartilaginous models of the digital bones). If interdigital cell death does not occur, a soft tissue web connects the digits on either side. There is more, however, to the development of digits than simply sculpting the interdigital spaces by cell death. A future digit is first recognizable as a longitudinal condensation of mesenchyme, which soon begins to lay down a precartilaginous matrix. Each digit develops its own character, as determined by the number of phalangeal segments or its specific size and shape. The identity of individual digits is not fixed until relatively late in limb formation. The exception to this rule is the first digit (thumb), which forms independently of shh. The identity of the remainder of the digits is determined by the concentration and the duration of exposure of their cells to shh. Digit 2 is formed from cells that have been exposed to shh, but have not themselves produced this signaling factor. A longer period of exposure and a greater concentration of shh is required to form digit 4, and digit 5 requires the longest exposure time and concentration of shh. Although the shh-based mechanism outlined above can account for their nature, it does not account for the number of digits. Increasing evidence suggests that a Turing* mechanism involving interactions between activating and inhibiting molecules might explain the basis for digit development. Whereas the Turing model * Alan Turing is best known as the father of modern computer science and artificial intelligence, but in 1952 he published a theoretical model for the formation of regular patterns from an initially homogeneous mix of activating and inhibiting substances. Although it took biologists many decades to appreciate this work, it now provides an explanation for the appearance of features as diverse as the stripes on zebras to the number of digits in a hand. In humans and many mammals, the normal number of digits is 5, but Hox gene deletion experiments have shown that when some are inactivated, extra digits form and the more Hox genes that are inactivated, the more digits form (as many as 14 in a single hand). All human digits contain three phalangeal segments except for the first digits (thumb and great toe), which consist of only two segments. The differentiation and histogenesis of the specific tissue components of the limb are later developmental events that build on the morphogenetic blueprint already established. Skeleton the skeleton is the first major tissue of the limb to show signs of overt differentiation. Its gross morphology, whether normal or abnormal, closely reflects the major pattern-forming events that shape the limb as a whole. Formation of the skeleton can be first seen as a condensation of mesenchymal cells in the central core of the proximal part of the limb bud. Even before undergoing condensation, these cells are determined to form cartilage, and if they are transplanted to other sites or into culture, they differentiate only into cartilage. The ectoderm of the limb bud exerts an inhibitory effect on cartilage differentiation, so cartilage does not form in the region just beneath the ectoderm. On the dorsal side of the limb bud, mesenchymal cells are prevented from differentiating into cartilage by Wnt-7a, produced by the ectoderm. While skeletal development continues, their expression is progressively restricted to the cells that become the perichondrium or periosteum surrounding the bones. Differentiation of the cartilaginous skeleton occurs in a proximodistal sequence, and in mammals the postaxial structures of the distal limb segments differentiate before the preaxial structures. The postaxial skeleton of the arm is considered to be the humerus, ulna, digits 2 through 5, and their corresponding carpal and metacarpal elements. The preaxial portion of the limb bud is progressively reduced during limb outgrowth and contributes only to the radius and possibly the first digital ray. The development of the limb girdles remains incompletely investigated, but experimental work on the chick has shown that the blade of the scapula is derived from cells of the dermomyotome, whereas the remainder of the scapula arises from lateral plate mesoderm. The three bones of the pelvis all arise from lateral plate mesoderm, with no known contribution from the somites. Each of the bones of the pelvis, as well as the two developmentally different components of the scapula, is characterized by a different molecular signature. How the bones of the appendages are patterned to connect with their respective girdles is still poorly understood, but studies of mutants suggest that the transcription factors Pbx-1 and Pbx-2 play an important upstream role. A characteristic feature in differentiation of the limb skeleton is the formation of joints. Joint formation occurs by the transverse splitting of precartilaginous rods, rather than by the apposition of two separate skeletal elements. At the site of a future joint, cells expressing Gdf-5 condense in the interzone and then begin to differentiate into the major tissues constituting the joint - articular cartilage, synovial membrane and intrajoint ligaments. It is presently not known whether the original interzone cells are homogeneous and later differentiate into the specific joint components or if the original interzone consists of a mixed population of cells already determined to form the different joint tissues. These latter phases of joint formation require movement generated by contractions of the muscles that traverse the joints. Although the early development of a joint is independent of movement, the absence of movement at later stages results in ankylosis or "freezing" of the joint. A well-known mutant family, called brachypodism, involves a shortening of the limb and the lack of development of certain joints, specifically the interphalangeal joints. There are five major groups of brachydactylies (shortening of the digits), each of which contains several subtypes. Musculature the musculature of the limb is derived from myogenic cells that migrate into the very early limb bud from the ventral part of the dermomyotome of the somite. Each somite in the limb region contributes 30 to 100 migratory precursor cells to the future limb musculature. These cells are stimulated to leave the somite and migrate toward the limb through the stimulus of scatter factor (hepatic growth factor), which is produced by the proximal cells of the limb-forming area. Before migrating, the premuscle cells in the somite express c-met, which is a specific receptor for scatter factor. The premyogenic cells, which are morphologically indistinguishable from the other mesenchymal cells, express Pax-3 and spread throughout the limb bud. In the splotch mutant, which is characterized by the absence of Pax-3 expression, muscle cells do not populate the limb bud. Migrating premuscle cells also express the cell adhesion molecule N-cadherin, which is important in correctly distributing them throughout the limb bud mesenchyme. The migrating premuscle cells keep pace with the elongation of the limb bud, although cells expressing characteristic muscle molecules. Some experimental evidence suggests that premyogenic cells are not present in the distal mesenchyme. Actual differentiation of the premuscle cells into muscle within the limb requires signals from the ectoderm, principally Wnt-6. If the ectoderm of the limb bud is removed, cartilage and connective tissue, but not muscle, differentiate. Shortly after the condensations of the skeletal elements take shape, the myogenic cells themselves begin to coalesce into two common muscle masses: one the precursor of the flexor muscles and the other giving rise to the extensor muscles. The transcription factor Tcf-4 is expressed throughout the connective tissue associated with the muscle masses. This is the connective tissue that determines the morphology of the individual muscles as they take shape. The next stage in muscle formation is the splitting of the common muscle masses into anatomically recognizable precursors of the definitive muscles of the limb. Little is known about the mechanisms that guide the splitting of the common muscle masses, although more recent evidence suggests that the pattern of blood vessels defines the future sites of cleavage through the secretion of platelet-derived growth factor and its action on the formation of connective tissue sheaths around the forming muscles. The fusion of myoblasts into early myotubes begins to occur during these early stages of muscle development. Considerable evidence suggests that myogenic precursor cells do not possess intrinsic information guiding their morphogenesis. Rather, the myogenic cells follow the lead of connective tissue cells, which are the bearers and effectors of the morphogenetic information required to form anatomically correct muscles. In experiments in which the somites normally associated with a limb bud are removed and replaced by somites from elsewhere along the body axis, myogenic cells are morphogenetically neutral. Muscle morphogenesis is typically normal even though the muscle fiber precursors are derived from abnormal sources. A later function of the T-box transcription factors Tbx-5 and Tbx-4, which play earlier important roles in initiating the development of the forelimbs and hindlimbs, respectively (see p. Depending on the specific muscle, the migration, fusion, or displacement of muscle primordia may be involved in the genesis of the final form of the muscle. In one case, genetically programmed cell death, apoptosis, is responsible for the disappearance of an entire muscle layer (the contrahentes muscle) in the flexor side of the human hand. The myogenic cells differentiate to the myotube stage; they then accumulate glycogen and soon degenerate. The reason it degenerates in the human hand at such a late stage in its differentiation is not understood. In (B) Gdf+ cells accumulating at the site of the future joint provide the cellular basis for most joint tissues. Myoblasts of the trapezius and sternocleidomastoid muscles (branchiomeric muscles) originate from lateral plate mesoderm at the occipital level. The deep shoulder girdle muscles (rhomboids and serratus anterior) are myotomally derived. The superficial shoulder muscles (pectoralis and latissimus dorsi) have a more complex origin. They arise from the pool of myogenic cells that originally migrated from the somite to the limb bud, but some of the descendants of this original pool later migrate from the common muscle masses in the limb back into the trunk, while retaining a cellular connection with the proximal humerus that will be their anatomical insertion. Although limb muscles assume their definitive form in the very early embryo, they must undergo considerable growth in length and cross-sectional area to keep up with the overall growth of the embryo. The added satellite cell nuclei increase the potential of the muscle fiber to produce structural and contractile proteins, which increase the cross-sectional area of each muscle fiber. Accompanying this addition to the nuclear complement of the muscle fibers is their lengthening by the addition of more sarcomeres, usually at the ends of the muscle fibers. Although the muscles are capable of contracting in the early fetal period, their physiological properties continue to mature until after birth. Tendons To function properly, muscles must attach to bones through the formation of tendons. A tendon is a band of dense fibrous connective tissue that is attached to the muscle through the myotendinous junction and to the bone through the enthesis, a complex structure with four zones forming a gradient from type I collagen to fibrocartilage and cartilage and, finally, an actual osseous union with the bone. Early experiments showed that when the somites adjoining the limb-forming regions were removed, the limbs developed without muscles, but rudimentary tendons did appear, although they later degenerated. These experiments showed that muscle fibers arise from somitic mesoderm, whereas limb tendons originate from lateral plate mesoderm. The tendons in limbs, axial structures, and the head require different conditions for their formation. Muscle is not needed for their early formation, but interactions with muscle are required for the later differentiation of tendons.
Review: Properties of sperm and seminal fluid erectile dysfunction lotion viagra jelly 100 mg buy overnight delivery, informed by research on reproduction and contraception impotence mayo clinic discount 100 mg viagra jelly mastercard. The role of molecular chaperones in spermatogenesis and the post-testicular maturation of mammalian spermatozoa erectile dysfunction va rating order viagra jelly 100 mg with visa. Recent advance in our understanding of the molecular nature of chromosomal abnormalities erectile dysfunction treatment aids viagra jelly 100 mg mastercard. The road to maturation somatic cell interaction and self-organization of the mammalian oocyte erectile dysfunction vasectomy discount viagra jelly 100 mg free shipping. Germ cellintrinsic and extrinsic factors govern meiotic initiation in mouse embryos. Activins, inhibins, and follistatins: further thoughts on a growing family of regulators. Molecular events and signalling pathways of male germ cell differentiation in mouse. The luteinizing hormone surge: the final stage in ovulation induction: modern aspects of ovulation triggering. Physiology and pathophysiology of the human spermatozoon: the role of electron microscopy. Shortly before ovulation, the epithelial cells of the uterine tube become more highly ciliated and smooth muscle activity in the tube and its suspensory ligament increases as the result of hormonal influences. By ovulation, the fimbriae of the uterine tube move closer to the ovary and seem to sweep rhythmically over its surface. This action, in addition to the currents set up by the cilia, efficiently captures the ovulated egg complex. Capture of the egg by the uterine tube also involves an adhesive interaction between,ihe egg complex and the ciliary surface of the tube. Even without these types of natural adaptations, the ability of the uterine tubes to capture eggs is remarkable. If the fimbriated end of tlie tube has een removed, egg capture occurs remarkably often, anci pregnancies have even occurred in women who have had one ovary ana the contralateral uterine tube removed. In such cases the ovulated egg would have to travel free in the pelvic cavity fo a considerable distance before entering the ostium of the uteri e tube on the other side. When inside the uterine tube, the egg is transported toward the uterus, mainly as the result of contractions of the smooth musculature of the tubal wall. Although the cilia lining the tubal mucosa may also play a role in egg transport, their action is not obligatory because women with immotile cilia syndrome are often fertile. While in the uterine tube, the egg is bathed in tubal fluid, which is a combination of secretion by the tubal epithelial cells and transudate from capillaries just below the epithelium. In some mammals, exposure to oviductal secretions is important to the survival of the ovum and for modifying the composition of the zona pellucida, but the role of tubal fluid in humans is less clear. Egg transport typically occurs in two phases: slow transport in the ampulla (approximately 72 hours) and a more rapid phase (8 hours) during which the egg or embryo passes through the isthmus and into the uterus (see p. By a poorly understood mechanism, possibly local edema or reduced muscular activity, the egg is temporarily prevented from entering the isthmic portion of the tube, but under the influence of progesterone, the uterotubal junction relaxes and permits entry of the ovum. By roughly 80 hours after ovulation, the ovulated egg or embryo has passed from the uterine tube into the uterus. It also describes the cyclic, hormonally controlled changes in the female reproductive tract that ready it for fertilization and the support of embryonic development. This article first explains how the egg and sperm cells come together in the female reproductive tract so that fertilization can occur. It then outlines the complex set of interactions involved in fertilization of the egg by a sperm. The first meiotic division is completed, and the second meiotic division proceeds until the metaphase stage, at which point the second meiotic arrest occurs. With the increased blood flow, plasma proteins leak into the tissues through the postcapillary venules, witli esulting local edema. The edema and the release of certain p armacologically active compounds, such as prostagla dins, histamine, vasopressin, and plasminogen activator, provide the starting point for a series of reactions that result in the local production of matrix metalloproteinases-a family of 1 $:tic enzymes that degrade components of the extracellular matrix. At the same time, the secretion of hyaluronic acid by cells of the cumulus results in a loosening of the cells surrounding the egg. Within minutes after rupture of the follicular wall, the cumulus oophorus detaches from the granulosa, and the egg is released from the ovary. Ovulation results in the expulsion of both antral fluid and the ovum from the ovary into the peritoneal cavity. The ovum is not ovulated as a single naked cell, but as a complex consisting of (1) the ovum, (2) the zona pellucida, (3) the two- to three-cell-thick corona radiata, and (4) a sticky matrix containing surrounding cells of the cumulus oophorus. By convention, the adhering cumulus cells are designated the corona radiata after ovulation has occurred. Often called mittelschmerz (German for "middle pain"), this pain may accompany slight bleeding from the ruptured follicle. Sperm Transport Sperm transport occurs in both the male reproductive tract and the female reproductive tract. In the male reproductive tract, transport of spermatozoa is closely connected with their structural and functional maturation, whereas in the female reproductive tract, it is important for spermatozoa to pass to the upper uterine tube where they can meet the ovulated egg. In the preovulatory follicle, the cumulus cells (arrow) are tightly packed around the oocyte. Spermatozoa are passively transported via testicular fluid from the seminiferous tubules to the caput (head) of the epididymis through the rete testis and the efferent ductules. They are propelled by fluid pressure generated in the seminiferous tubules and through contractions of myoid cells that lie outside the seminiferous tubules. Their progress is then assisted by smooth muscle contractions and ciliary currents in the efferent ductules. Spermatozoa spend approximately 12 days in the highly convoluted duct of the epididymis, which measures 6 m in the human, during which they undergo biochemical maturation (Box 2. This period of maturation is associated with changes in the glycoproteins in the plasma membrane of the sperm head. By the time the spermatozoa have reached the cauda (tail) of the epididymis, they are capable of fertilizing an egg. On ejaculation, the spermatozoa rapidly pass through the ductus deferens and are mixed with fluid secretions from the seminal vesicles and prostate gland. Prostatic fluid is rich in citric acid, acid phosphatase, zinc, and magnesium ions, whereas fluid of the seminal vesicle is rich in fructose (the principal energy source of spermatozoa) and prostaglandins. The 2 to 6 mL of ejaculate (semen, or seminal fluid) typically consists of 40 to 250 million spermatozoa mixed with alkaline fluid from the seminal vesicles (approximately 70% of the total) and acid secretion (pH, 6. Despite the numerous spermatozoa (>100 million) normally present in an ejaculate, a number as small as 25 million spermatozoa per ejaculate may be compatible with fertility. In the female reproductive tract, sperm transport begins in the upper vagina and ends in the ampulla of the uterine tube, where the spermatozoa make contact with the ovulated egg. In (B) numbers of spermatozoa typically found in various parts of the female reproductive tract are indicated in red. The acidic vaginal fluid normally serves a bactericidal function in protecting the cervical canal from pathogenic organisms. The buffering effect lasts only a few minutes in humans, but it is enough time for the spermatozoa to approach the cervix in an environment (pH, 6. Within a minute after entry into the vagina, human semen coagulates into a loose gel. The coagulum protects the spermatozoa from the harsh vaginal microenvironment and may also hold the sperm close to the cervical os. A major factor in generating the seminal gel is the presence of semenogelin proteins contributed by the seminal vesicles. The next barriers that the sperm cells must overcome are the cervical canal and the cervical mucus that blocks it. Swimming movements by the spermatozoa are important for the penetration of the cervical mucus. The composition and viscosity of cervical mucus vary considerably throughout the menstrual cycle. Composed of cervical mucin (a glycoprotein with a high carbohydrate composition) and soluble components, cervical mucus is not readily penetrable. Between days 9 and 16 of the cycle, however, its water content increases, and this change facilitates the passage of sperm through the cervix around the time of ovulation; such mucus is sometimes called E mucus. After ovulation, under the influence of progesterone, the production of watery cervical mucus ceases, and a new type of sticky mucus, which has a much-decreased water content, is produced. This progestational mucus, sometimes called G mucus, is almost completely resistant to sperm penetration. A highly effective method of natural family planning makes use of the properties of cervical mucus. In addition to its low pH, the vagina and also the cervix use a variety of immunological strategies to prevent pathogens from entering the upper reaches of the female reproductive tract. These include cells (mainly neutrophils and macrophages), immunoglobulins (mainly IgA and IgG), and complement proteins to protect against invaders. Although spermatozoa do stimulate some of these defenses, they are able to avoid most of them in their transit into and through the cervix. Nevertheless, after 24 hours, phagocytosis of spermatozoa seriously reduces the number of viable spermatozoa that could potentially fertilize an egg. Some studies have suggested the presence of cervical crypts, which can potentially hold viable sperm for several days. Others have shown the presence of small cervical grooves or canals that could guide spermatozoa in their transit through the cervical canal. Linear strands of cervical mucus may also play a role in the cervical transport of spermatozoa. Such rapid transport relies more on muscular movements of the female reproductive tract than on the motility of the spermatozoa themselves. These early arriving sperm, however, appear not to be as capable of fertilizing an egg as do those that have spent more time in the female reproductive tract. The second, slow phase of sperm transport involves the swimming of spermatozoa through the cervical mucus (traveling at a rate of 2 to 3 mm/h), their storage in cervical crypts, and their final passage through the cervical canal as much as 2 to 4 days later. According to some calculations, swimming movements alone could drive spermatozoa across the uterine cavity in slightly more than 10 minutes, but contractions of the uterine musculature play an important role. Late in the follicular phase of the menstrual cycle, cranially directed waves of smooth muscle contractions in the uterine wall increase in intensity, and these can propel spermatozoa toward the uterotubal junction. Increasing evidence exists that the ovary containing the dominant follicle that will be ovulated influences the uterine tube on that side to serve as a preferential passageway for sperm. A complex ovarian circulatory pathway provides a pathway for carrying hormones emanating from the dominant ovary to the same side of the uterus. In addition, the temperature of the uterine tube on that side has been shown to be 1. These influences result in a greater patency of the ipsilateral uterotubular junction than the one leading from the tube that will not carry the ovulated egg. According to some estimates, only several hundred spermatozoa enter the uterine tubes, and most enter the tube containing the ovulated egg. In mice, calmegin, a chaperone protein on sperm, is required for sperm to enter the uterotubal junction. This suggests the presence of a molecular recognition system that regulates entry of sperm into the uterine tube. Once inside the uterine tube, the spermatozoa collect in the isthmus and bind to the epithelium for approximately 24 hours. During this time, they are influenced by secretions of the tube to undergo the capacitation reaction. One phase of capacitation is removal of cholesterol from the surface of the sperm. The next phase of capacitation consists of removal of many of the glycoproteins that were deposited on the surface of the spermatozoa during their tenure in the epididymis. Capacitation is required for spermatozoa to be able to fertilize an egg (specifically, to undergo the acrosome reaction; see p. After the capacitation reaction, the spermatozoa undergo a period of hyperactivity and detach from the tubal epithelium. Hyperactivation helps the spermatozoa break free of the bonds that held them to the tubal epithelium. It also assists the sperm in penetrating isthmic mucus and the corona radiata and the zona pellucida, which surround the ovum. On their release from the isthmus, the spermatozoa make their way up the tube through a combination of muscular movements of the tube and swimming movements. The simultaneous transport of an egg down and spermatozoa up the tube is currently explained on the basis of peristaltic contractions of the uterine tube muscles. Within a given compartment, the gametes are caught up in churning movements that during 1 or 2 days bring the egg and spermatozoa together. Fertilization of the egg normally occurs in the ampullary portion (upper third) of the uterine tube. Estimates suggest that spermatozoa retain their function in the female reproductive tract for approximately 80 hours. Some spermatozoa actually pass through the fimbriated end of the uterine tube and make their way into the peritoneal cavity. In these cases, fertilization likely occurred through sperm passing through the contralateral patent uterine tube and into the peritoneal cavity. From there, they would have been picked up by the ampullary end of the blocked tube and carried down to the ovulated egg, where fertilization occurred.
References
- Kader A, Frazzini V, Baker C, et al: Effect of mild hypothermia on nitric oxide synthesis during focal cerebral ischemia, Neurosurgery 35:272-277, 1994.
- Brauer RO. Repair of unilateral cleft lip. Triangular flap repairs. Clin Plast Surg 1985;12:595.
- Keszler M, Subramanian KN, Smith YA, Dhanireddy R, Mehta N, Molina B, Cox CB, Moront MG. Pulmonary Management During Extracorporeal Membrane Oxygenation. Critical Care Medicine 1989;17:495-500.
- Suchdev, P.S., et al., Overview of the Biomarkers Reflecting Inflammation and Nutritional Determinants of Anemia (BRINDA) Project. Adv. Nutr., 2016.
- Angelov L, Mohammadi AM, Bennett EE, et al. Impact of 2-staged stereotactic radiosurgery for treatment of brain metastases ? 2 cm. J Neurosurg 2017:1-17.
- Kawai Y, Tsukamoto E, Nozaki Y, et al: Signifance of reduced uptake of iodinated fatty acid analogue for the evaluation of patients with acute chest pain. J Am Coll Cardiol 2001;38:1888-1894.